
Reviews: Immunity To Fungal Infections
Reviews: Immunity To Fungal Infections
You might also like
- Compensatory MechanismDocument30 pagesCompensatory MechanismSenthil Kumar Elumalai100% (1)
- Senses Review WorksheetDocument6 pagesSenses Review WorksheetpicassaaNo ratings yet
- 008 GB e Vidas Assay SolutionsDocument4 pages008 GB e Vidas Assay SolutionsvishnupgiNo ratings yet
- Blrincon - Blrincon - 2023 Nature ReviewsDocument20 pagesBlrincon - Blrincon - 2023 Nature ReviewsDavidf VillabonaNo ratings yet
- Inmunidad de Las Infecciones FungicasDocument14 pagesInmunidad de Las Infecciones FungicasGabriel Gonzalez BinottoNo ratings yet
- Ni Hms 316844Document15 pagesNi Hms 316844ntnquynhproNo ratings yet
- Immune Response To Fungal Infections: Jose L. Blanco, Marta E. GarciaDocument24 pagesImmune Response To Fungal Infections: Jose L. Blanco, Marta E. Garciagagah152No ratings yet
- Immunity Against Fungi: Michail S. Lionakis, Iliyan D. Iliev, and Tobias M. HohlDocument17 pagesImmunity Against Fungi: Michail S. Lionakis, Iliyan D. Iliev, and Tobias M. HohlFajar SukmajayaNo ratings yet
- Allergy and Immunity To Fungal Infections and Colonization - European Respiratory SocietyDocument17 pagesAllergy and Immunity To Fungal Infections and Colonization - European Respiratory SocietyMuhammed SabdatNo ratings yet
- Pathogenic Fungal Infection in The Lung: Zhi Li, Gen Lu and Guangxun MengDocument20 pagesPathogenic Fungal Infection in The Lung: Zhi Li, Gen Lu and Guangxun Mengbanteng wibisonoNo ratings yet
- Co-Infection Subverts Mucosal Immunity in The Upper Respiratory TractDocument7 pagesCo-Infection Subverts Mucosal Immunity in The Upper Respiratory TractHarold Andres Ortiz CortesNo ratings yet
- AntivirL B y T en PulmonDocument9 pagesAntivirL B y T en PulmonAnailil MoralesNo ratings yet
- Biomedicine & Pharmacotherapy: ReviewDocument12 pagesBiomedicine & Pharmacotherapy: Reviewkorede juniorNo ratings yet
- Immunotherapy A Potential Adjunctive Treatment For Candida - Il e IfnDocument6 pagesImmunotherapy A Potential Adjunctive Treatment For Candida - Il e IfnLuiz JmnNo ratings yet
- Antifungal Drugs - The Current Armamentarium and Development of New AgentsDocument20 pagesAntifungal Drugs - The Current Armamentarium and Development of New AgentssovalaxNo ratings yet
- Reviews: Modulating Immunity As A Therapy For Bacterial InfectionsDocument12 pagesReviews: Modulating Immunity As A Therapy For Bacterial InfectionsZineb RAISNo ratings yet
- Immunity Against Fungal Infections: Shuai JiangDocument4 pagesImmunity Against Fungal Infections: Shuai JiangShafira AzzahraNo ratings yet
- Fungal Infections and Critically Ill AdultsDocument6 pagesFungal Infections and Critically Ill AdultsBejinaruOanaSiCristiNo ratings yet
- MICP Bacterial Infections s1Document12 pagesMICP Bacterial Infections s1Valerian VolkzkiNo ratings yet
- Antimicrobial Resistance in Hospital-Acquired Gram-Negative Bacterial InfectionsDocument9 pagesAntimicrobial Resistance in Hospital-Acquired Gram-Negative Bacterial InfectionsAlina BanciuNo ratings yet
- Therapy by HelminthsDocument10 pagesTherapy by HelminthsZineb RAISNo ratings yet
- Fungal Infections in The IcuDocument23 pagesFungal Infections in The IcuVemuri SrinivasNo ratings yet
- And Antibacterial Resistance: Pseudomonas Aeruginosa: Burn Infection, TreatmentDocument10 pagesAnd Antibacterial Resistance: Pseudomonas Aeruginosa: Burn Infection, TreatmentnurelisabayuNo ratings yet
- Immunity To Fungi: Tobias M Hohl, Amariliz Rivera and Eric G PamerDocument8 pagesImmunity To Fungi: Tobias M Hohl, Amariliz Rivera and Eric G PamerPieralessandro LasalviaNo ratings yet
- A Molecular Perspective of Microbial Pathogenicity PDFDocument11 pagesA Molecular Perspective of Microbial Pathogenicity PDFjon diazNo ratings yet
- TUBERCULOSISDocument13 pagesTUBERCULOSISSam VattaraiNo ratings yet
- Innate and Adaptive Immune ResponseDocument12 pagesInnate and Adaptive Immune ResponseMaulidza SiltaNo ratings yet
- Microbiology PointsDocument17 pagesMicrobiology Points1031 Muhammad zaryabNo ratings yet
- Immunology of MycobacteriumDocument37 pagesImmunology of MycobacteriumPhablo vinicius dos santos carneiroNo ratings yet
- Immunological, Cellular and Molecular Events in Typhoid FeverDocument12 pagesImmunological, Cellular and Molecular Events in Typhoid FeverAde RifkaNo ratings yet
- Propionibacterium Acnes: A Putative Immunemodulating Weapon Against The Coronavirus Impending EpidemyDocument10 pagesPropionibacterium Acnes: A Putative Immunemodulating Weapon Against The Coronavirus Impending EpidemyInternational Journal of Innovative Science and Research TechnologyNo ratings yet
- Plasmodium Infection and Drug Cure For Malaria Vaccine DevelopmentDocument22 pagesPlasmodium Infection and Drug Cure For Malaria Vaccine DevelopmentZowonu Prince HorlortuNo ratings yet
- Avanços Vacinas Contra Fungos 2007Document33 pagesAvanços Vacinas Contra Fungos 2007Bruno Nogueira BarrosNo ratings yet
- The Role of Fungi in Diseases of The Nose and Sinuses: NARAC ReviewDocument8 pagesThe Role of Fungi in Diseases of The Nose and Sinuses: NARAC ReviewPinandhito LatukolanNo ratings yet
- Finals Mycobacteria 2Document14 pagesFinals Mycobacteria 2JaneNo ratings yet
- Ajidm 1 4 2Document6 pagesAjidm 1 4 2rehanaNo ratings yet
- Seminario ArtículoDocument20 pagesSeminario Artículomarixa ledesmaNo ratings yet
- Medical Surgical Nursing Module 2Document6 pagesMedical Surgical Nursing Module 2JAY TOM BOLIVARNo ratings yet
- GetahunLTBINEJM2015 PDFDocument9 pagesGetahunLTBINEJM2015 PDFSharah Stephanie IINo ratings yet
- Mucormycosis in Immunocompetent Patients - A Case-Series of Patients With Maxillary Sinus Involvement and A Critical Review of The LiteratureDocument8 pagesMucormycosis in Immunocompetent Patients - A Case-Series of Patients With Maxillary Sinus Involvement and A Critical Review of The LiteratureAnoop SinghNo ratings yet
- Viral Structure NotesDocument9 pagesViral Structure Notesantonia putriNo ratings yet
- Mico PlasmasDocument11 pagesMico PlasmasZuri JiménezNo ratings yet
- Iran Red Crescent Med J 2009 11 3 244Document10 pagesIran Red Crescent Med J 2009 11 3 244rmkamundimuNo ratings yet
- Pseudomonas Baru 4-1Document29 pagesPseudomonas Baru 4-1Nadia Iriana DewiNo ratings yet
- Myco Viro Notes Module 6Document6 pagesMyco Viro Notes Module 6Kyle Jude Roberts SanesNo ratings yet
- Fungal Infections of The Gastrointestinal TractDocument30 pagesFungal Infections of The Gastrointestinal Tractope kolinaNo ratings yet
- Helmintos NematodosDocument12 pagesHelmintos NematodosHANNIANo ratings yet
- Yersinia InmunidadDocument12 pagesYersinia InmunidadJ Carlos MorenoNo ratings yet
- Fungal Infections in Diabetes Mellitus: An Overview: Review ArticleDocument5 pagesFungal Infections in Diabetes Mellitus: An Overview: Review ArticleAudrey Ira YunitaNo ratings yet
- Wa0006.Document12 pagesWa0006.Jose VillasmilNo ratings yet
- Introduction To Medical MicrobiologyDocument9 pagesIntroduction To Medical MicrobiologyIsba Shadai Estrada GarciaNo ratings yet
- Key Points Revision: 1 Structure and Classification of BacteriaDocument17 pagesKey Points Revision: 1 Structure and Classification of Bacteriatarun956519No ratings yet
- CandidaDocument33 pagesCandidaAlberto ValenzuelaNo ratings yet
- EBS ProjectDocument9 pagesEBS Projectdiksha halderNo ratings yet
- Mycology - Chapter Seven Opportunistic Mycoses: Let Us Know What You ThinkDocument9 pagesMycology - Chapter Seven Opportunistic Mycoses: Let Us Know What You ThinkAreeqa AliNo ratings yet
- Pathogenesis of Fungal InfectionsDocument70 pagesPathogenesis of Fungal Infectionsማላያላም ማላያላም100% (15)
- Covid 19 ReviewDocument22 pagesCovid 19 ReviewQueencyNo ratings yet
- Module # 6-Student Activity Sheet - Opportunistic MycosesDocument9 pagesModule # 6-Student Activity Sheet - Opportunistic MycosesYlia MastarsNo ratings yet
- Mucormycosis: Continuing Education ActivityDocument5 pagesMucormycosis: Continuing Education ActivityVarun kkNo ratings yet
- Identification and Construction of A Multi Epitopes Vaccine Design Against Klebsiella Aerogenes: Molecular Modeling StudyDocument16 pagesIdentification and Construction of A Multi Epitopes Vaccine Design Against Klebsiella Aerogenes: Molecular Modeling StudySamer ShamshadNo ratings yet
- Finals MycobacteriaDocument15 pagesFinals MycobacteriaJaneNo ratings yet
- Histoplasmosis AtsDocument4 pagesHistoplasmosis Atsanabella081096No ratings yet
- Management of Infections in the Immunocompromised HostFrom EverandManagement of Infections in the Immunocompromised HostBrahm H. SegalNo ratings yet
- Lavees Cosmedics: Cara PakaiDocument2 pagesLavees Cosmedics: Cara PakaimonitamiftahNo ratings yet
- Oral Collagen Supplementation For Skin Aging: A Fad or The Future?Document3 pagesOral Collagen Supplementation For Skin Aging: A Fad or The Future?monitamiftahNo ratings yet
- Asam Traneksamat Untuk KulitDocument2 pagesAsam Traneksamat Untuk KulitmonitamiftahNo ratings yet
- SunscreenDocument8 pagesSunscreenmonitamiftahNo ratings yet
- Seminar: Heidi D NelsonDocument11 pagesSeminar: Heidi D NelsonmonitamiftahNo ratings yet
- LaserDocument104 pagesLasermonitamiftah100% (1)
- Lehninger Principles of Biochemistry PDFDocument37 pagesLehninger Principles of Biochemistry PDFmonitamiftah100% (1)
- Apoptosis & Nekrosis: Monita Sugianto Dosen Pengampu: Edhyana K. Sahiratmadja, DR., PHDDocument34 pagesApoptosis & Nekrosis: Monita Sugianto Dosen Pengampu: Edhyana K. Sahiratmadja, DR., PHDmonitamiftahNo ratings yet
- Guidelines Acne Vulgaris PDFDocument14 pagesGuidelines Acne Vulgaris PDFmonitamiftahNo ratings yet
- Apoptosis & Nekrosis: Monita Sugianto Dosen Pengampu: Edhyana K. Sahiratmadja, DR., PHDDocument34 pagesApoptosis & Nekrosis: Monita Sugianto Dosen Pengampu: Edhyana K. Sahiratmadja, DR., PHDmonitamiftahNo ratings yet
- Inflammation in AgingDocument14 pagesInflammation in AgingmonitamiftahNo ratings yet
- Efek Immunologi Radiasi Sinar UV MatahariDocument16 pagesEfek Immunologi Radiasi Sinar UV MataharimonitamiftahNo ratings yet
- Management of Aging SkinDocument19 pagesManagement of Aging SkinmonitamiftahNo ratings yet
- Efek Immunologi Radiasi Sinar UV MatahariDocument16 pagesEfek Immunologi Radiasi Sinar UV MataharimonitamiftahNo ratings yet
- Chapter 027Document3 pagesChapter 027barbaraNo ratings yet
- ALBCchicken Assessment 2Document6 pagesALBCchicken Assessment 2Filip Marian100% (1)
- Quick Quiz! 7-5: CHAPTER 7 Miscellaneous ProtozoaDocument17 pagesQuick Quiz! 7-5: CHAPTER 7 Miscellaneous ProtozoaTawfeeq AuqbiNo ratings yet
- Rash FeverDocument1 pageRash FeverSyima MnnNo ratings yet
- Farmacología de ItraconazolDocument5 pagesFarmacología de ItraconazolalexqaNo ratings yet
- Anatomy Physiology of The BrainDocument107 pagesAnatomy Physiology of The BrainRadha RamineniNo ratings yet
- Extraordinary Master VesselsDocument11 pagesExtraordinary Master VesselssillymindsNo ratings yet
- Exclusive Sellers of Chanson Bleu ChartreuxDocument9 pagesExclusive Sellers of Chanson Bleu ChartreuxpsykosomatikNo ratings yet
- Activity 8.2.4 Brain Anatomy Answer Key: Parietal LobeDocument5 pagesActivity 8.2.4 Brain Anatomy Answer Key: Parietal LobeAlexis CarranzaNo ratings yet
- Tài liệu học thêm 12A4 Dop: 15/3/2021 Revision (Vocabualary) I.Review 1.1 .Read the passageDocument10 pagesTài liệu học thêm 12A4 Dop: 15/3/2021 Revision (Vocabualary) I.Review 1.1 .Read the passageNguyen Thi Phuong ThaoNo ratings yet
- Solutions For Genetics Problems - Bil 151: Parental Genotypes: Male: FemaleDocument10 pagesSolutions For Genetics Problems - Bil 151: Parental Genotypes: Male: FemaleLevy Necesito0% (1)
- Usbn Bahasa Inggris Sma It Nurul Muhajirin Kotanegara Sungkai UtaraDocument5 pagesUsbn Bahasa Inggris Sma It Nurul Muhajirin Kotanegara Sungkai UtaraNuryani AhmadNo ratings yet
- Diagnosis of Mycotoxicoses in Veterinary MedicineDocument10 pagesDiagnosis of Mycotoxicoses in Veterinary MedicineDamir SuljevicNo ratings yet
- Care of Patients With Chest TubesDocument2 pagesCare of Patients With Chest Tubesaurezea100% (1)
- A Dictionary of Every Day WantsDocument571 pagesA Dictionary of Every Day Wantsdonald1976No ratings yet
- Paediatrics State Exam QuestionsDocument4 pagesPaediatrics State Exam Questionsnasibdin100% (1)
- Urine Procedure WordDocument27 pagesUrine Procedure WordYashly VargheseNo ratings yet
- 2007 Skeletal Inventory Forms (Complete)Document15 pages2007 Skeletal Inventory Forms (Complete)Sergio F Cano100% (1)
- Cva CPDocument53 pagesCva CPApol Pen100% (2)
- 1.andersen: 2. BimlerDocument6 pages1.andersen: 2. BimlerRoxana ElenaNo ratings yet
- Anal Fissure1Document3 pagesAnal Fissure1Jehanzeb ZebNo ratings yet
- BLOODDocument1 pageBLOODJuana IlaganNo ratings yet
- Wirth (2016) Core Stability in Athletes A Critical Analysis of Current PDFDocument14 pagesWirth (2016) Core Stability in Athletes A Critical Analysis of Current PDFBiology PageNo ratings yet
- Pneumocystis PneumoniaDocument5 pagesPneumocystis Pneumoniadger11No ratings yet
- Protest Petition For Establishment of KVIC National Head Quarter in Capital of India - Abhishek KadyanDocument52 pagesProtest Petition For Establishment of KVIC National Head Quarter in Capital of India - Abhishek KadyanNaresh KadyanNo ratings yet
- Electrosurgery in Periodontics PerioDocument29 pagesElectrosurgery in Periodontics PerioFourthMolar.com100% (1)
- Karya Tulis IlmiahDocument50 pagesKarya Tulis IlmiahSayyid SultanNo ratings yet






































































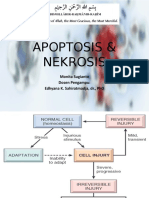
































Download as pdf or txt
You might also like
- Compensatory MechanismDocument30 pagesCompensatory MechanismSenthil Kumar Elumalai100% (1)
- Senses Review WorksheetDocument6 pagesSenses Review WorksheetpicassaaNo ratings yet
- 008 GB e Vidas Assay SolutionsDocument4 pages008 GB e Vidas Assay SolutionsvishnupgiNo ratings yet
- Blrincon - Blrincon - 2023 Nature ReviewsDocument20 pagesBlrincon - Blrincon - 2023 Nature ReviewsDavidf VillabonaNo ratings yet
- Inmunidad de Las Infecciones FungicasDocument14 pagesInmunidad de Las Infecciones FungicasGabriel Gonzalez BinottoNo ratings yet
- Ni Hms 316844Document15 pagesNi Hms 316844ntnquynhproNo ratings yet
- Immune Response To Fungal Infections: Jose L. Blanco, Marta E. GarciaDocument24 pagesImmune Response To Fungal Infections: Jose L. Blanco, Marta E. Garciagagah152No ratings yet
- Immunity Against Fungi: Michail S. Lionakis, Iliyan D. Iliev, and Tobias M. HohlDocument17 pagesImmunity Against Fungi: Michail S. Lionakis, Iliyan D. Iliev, and Tobias M. HohlFajar SukmajayaNo ratings yet
- Allergy and Immunity To Fungal Infections and Colonization - European Respiratory SocietyDocument17 pagesAllergy and Immunity To Fungal Infections and Colonization - European Respiratory SocietyMuhammed SabdatNo ratings yet
- Pathogenic Fungal Infection in The Lung: Zhi Li, Gen Lu and Guangxun MengDocument20 pagesPathogenic Fungal Infection in The Lung: Zhi Li, Gen Lu and Guangxun Mengbanteng wibisonoNo ratings yet
- Co-Infection Subverts Mucosal Immunity in The Upper Respiratory TractDocument7 pagesCo-Infection Subverts Mucosal Immunity in The Upper Respiratory TractHarold Andres Ortiz CortesNo ratings yet
- AntivirL B y T en PulmonDocument9 pagesAntivirL B y T en PulmonAnailil MoralesNo ratings yet
- Biomedicine & Pharmacotherapy: ReviewDocument12 pagesBiomedicine & Pharmacotherapy: Reviewkorede juniorNo ratings yet
- Immunotherapy A Potential Adjunctive Treatment For Candida - Il e IfnDocument6 pagesImmunotherapy A Potential Adjunctive Treatment For Candida - Il e IfnLuiz JmnNo ratings yet
- Antifungal Drugs - The Current Armamentarium and Development of New AgentsDocument20 pagesAntifungal Drugs - The Current Armamentarium and Development of New AgentssovalaxNo ratings yet
- Reviews: Modulating Immunity As A Therapy For Bacterial InfectionsDocument12 pagesReviews: Modulating Immunity As A Therapy For Bacterial InfectionsZineb RAISNo ratings yet
- Immunity Against Fungal Infections: Shuai JiangDocument4 pagesImmunity Against Fungal Infections: Shuai JiangShafira AzzahraNo ratings yet
- Fungal Infections and Critically Ill AdultsDocument6 pagesFungal Infections and Critically Ill AdultsBejinaruOanaSiCristiNo ratings yet
- MICP Bacterial Infections s1Document12 pagesMICP Bacterial Infections s1Valerian VolkzkiNo ratings yet
- Antimicrobial Resistance in Hospital-Acquired Gram-Negative Bacterial InfectionsDocument9 pagesAntimicrobial Resistance in Hospital-Acquired Gram-Negative Bacterial InfectionsAlina BanciuNo ratings yet
- Therapy by HelminthsDocument10 pagesTherapy by HelminthsZineb RAISNo ratings yet
- Fungal Infections in The IcuDocument23 pagesFungal Infections in The IcuVemuri SrinivasNo ratings yet
- And Antibacterial Resistance: Pseudomonas Aeruginosa: Burn Infection, TreatmentDocument10 pagesAnd Antibacterial Resistance: Pseudomonas Aeruginosa: Burn Infection, TreatmentnurelisabayuNo ratings yet
- Immunity To Fungi: Tobias M Hohl, Amariliz Rivera and Eric G PamerDocument8 pagesImmunity To Fungi: Tobias M Hohl, Amariliz Rivera and Eric G PamerPieralessandro LasalviaNo ratings yet
- A Molecular Perspective of Microbial Pathogenicity PDFDocument11 pagesA Molecular Perspective of Microbial Pathogenicity PDFjon diazNo ratings yet
- TUBERCULOSISDocument13 pagesTUBERCULOSISSam VattaraiNo ratings yet
- Innate and Adaptive Immune ResponseDocument12 pagesInnate and Adaptive Immune ResponseMaulidza SiltaNo ratings yet
- Microbiology PointsDocument17 pagesMicrobiology Points1031 Muhammad zaryabNo ratings yet
- Immunology of MycobacteriumDocument37 pagesImmunology of MycobacteriumPhablo vinicius dos santos carneiroNo ratings yet
- Immunological, Cellular and Molecular Events in Typhoid FeverDocument12 pagesImmunological, Cellular and Molecular Events in Typhoid FeverAde RifkaNo ratings yet
- Propionibacterium Acnes: A Putative Immunemodulating Weapon Against The Coronavirus Impending EpidemyDocument10 pagesPropionibacterium Acnes: A Putative Immunemodulating Weapon Against The Coronavirus Impending EpidemyInternational Journal of Innovative Science and Research TechnologyNo ratings yet
- Plasmodium Infection and Drug Cure For Malaria Vaccine DevelopmentDocument22 pagesPlasmodium Infection and Drug Cure For Malaria Vaccine DevelopmentZowonu Prince HorlortuNo ratings yet
- Avanços Vacinas Contra Fungos 2007Document33 pagesAvanços Vacinas Contra Fungos 2007Bruno Nogueira BarrosNo ratings yet
- The Role of Fungi in Diseases of The Nose and Sinuses: NARAC ReviewDocument8 pagesThe Role of Fungi in Diseases of The Nose and Sinuses: NARAC ReviewPinandhito LatukolanNo ratings yet
- Finals Mycobacteria 2Document14 pagesFinals Mycobacteria 2JaneNo ratings yet
- Ajidm 1 4 2Document6 pagesAjidm 1 4 2rehanaNo ratings yet
- Seminario ArtículoDocument20 pagesSeminario Artículomarixa ledesmaNo ratings yet
- Medical Surgical Nursing Module 2Document6 pagesMedical Surgical Nursing Module 2JAY TOM BOLIVARNo ratings yet
- GetahunLTBINEJM2015 PDFDocument9 pagesGetahunLTBINEJM2015 PDFSharah Stephanie IINo ratings yet
- Mucormycosis in Immunocompetent Patients - A Case-Series of Patients With Maxillary Sinus Involvement and A Critical Review of The LiteratureDocument8 pagesMucormycosis in Immunocompetent Patients - A Case-Series of Patients With Maxillary Sinus Involvement and A Critical Review of The LiteratureAnoop SinghNo ratings yet
- Viral Structure NotesDocument9 pagesViral Structure Notesantonia putriNo ratings yet
- Mico PlasmasDocument11 pagesMico PlasmasZuri JiménezNo ratings yet
- Iran Red Crescent Med J 2009 11 3 244Document10 pagesIran Red Crescent Med J 2009 11 3 244rmkamundimuNo ratings yet
- Pseudomonas Baru 4-1Document29 pagesPseudomonas Baru 4-1Nadia Iriana DewiNo ratings yet
- Myco Viro Notes Module 6Document6 pagesMyco Viro Notes Module 6Kyle Jude Roberts SanesNo ratings yet
- Fungal Infections of The Gastrointestinal TractDocument30 pagesFungal Infections of The Gastrointestinal Tractope kolinaNo ratings yet
- Helmintos NematodosDocument12 pagesHelmintos NematodosHANNIANo ratings yet
- Yersinia InmunidadDocument12 pagesYersinia InmunidadJ Carlos MorenoNo ratings yet
- Fungal Infections in Diabetes Mellitus: An Overview: Review ArticleDocument5 pagesFungal Infections in Diabetes Mellitus: An Overview: Review ArticleAudrey Ira YunitaNo ratings yet
- Wa0006.Document12 pagesWa0006.Jose VillasmilNo ratings yet
- Introduction To Medical MicrobiologyDocument9 pagesIntroduction To Medical MicrobiologyIsba Shadai Estrada GarciaNo ratings yet
- Key Points Revision: 1 Structure and Classification of BacteriaDocument17 pagesKey Points Revision: 1 Structure and Classification of Bacteriatarun956519No ratings yet
- CandidaDocument33 pagesCandidaAlberto ValenzuelaNo ratings yet
- EBS ProjectDocument9 pagesEBS Projectdiksha halderNo ratings yet
- Mycology - Chapter Seven Opportunistic Mycoses: Let Us Know What You ThinkDocument9 pagesMycology - Chapter Seven Opportunistic Mycoses: Let Us Know What You ThinkAreeqa AliNo ratings yet
- Pathogenesis of Fungal InfectionsDocument70 pagesPathogenesis of Fungal Infectionsማላያላም ማላያላም100% (15)
- Covid 19 ReviewDocument22 pagesCovid 19 ReviewQueencyNo ratings yet
- Module # 6-Student Activity Sheet - Opportunistic MycosesDocument9 pagesModule # 6-Student Activity Sheet - Opportunistic MycosesYlia MastarsNo ratings yet
- Mucormycosis: Continuing Education ActivityDocument5 pagesMucormycosis: Continuing Education ActivityVarun kkNo ratings yet
- Identification and Construction of A Multi Epitopes Vaccine Design Against Klebsiella Aerogenes: Molecular Modeling StudyDocument16 pagesIdentification and Construction of A Multi Epitopes Vaccine Design Against Klebsiella Aerogenes: Molecular Modeling StudySamer ShamshadNo ratings yet
- Finals MycobacteriaDocument15 pagesFinals MycobacteriaJaneNo ratings yet
- Histoplasmosis AtsDocument4 pagesHistoplasmosis Atsanabella081096No ratings yet
- Management of Infections in the Immunocompromised HostFrom EverandManagement of Infections in the Immunocompromised HostBrahm H. SegalNo ratings yet
- Lavees Cosmedics: Cara PakaiDocument2 pagesLavees Cosmedics: Cara PakaimonitamiftahNo ratings yet
- Oral Collagen Supplementation For Skin Aging: A Fad or The Future?Document3 pagesOral Collagen Supplementation For Skin Aging: A Fad or The Future?monitamiftahNo ratings yet
- Asam Traneksamat Untuk KulitDocument2 pagesAsam Traneksamat Untuk KulitmonitamiftahNo ratings yet
- SunscreenDocument8 pagesSunscreenmonitamiftahNo ratings yet
- Seminar: Heidi D NelsonDocument11 pagesSeminar: Heidi D NelsonmonitamiftahNo ratings yet
- LaserDocument104 pagesLasermonitamiftah100% (1)
- Lehninger Principles of Biochemistry PDFDocument37 pagesLehninger Principles of Biochemistry PDFmonitamiftah100% (1)
- Apoptosis & Nekrosis: Monita Sugianto Dosen Pengampu: Edhyana K. Sahiratmadja, DR., PHDDocument34 pagesApoptosis & Nekrosis: Monita Sugianto Dosen Pengampu: Edhyana K. Sahiratmadja, DR., PHDmonitamiftahNo ratings yet
- Guidelines Acne Vulgaris PDFDocument14 pagesGuidelines Acne Vulgaris PDFmonitamiftahNo ratings yet
- Apoptosis & Nekrosis: Monita Sugianto Dosen Pengampu: Edhyana K. Sahiratmadja, DR., PHDDocument34 pagesApoptosis & Nekrosis: Monita Sugianto Dosen Pengampu: Edhyana K. Sahiratmadja, DR., PHDmonitamiftahNo ratings yet
- Inflammation in AgingDocument14 pagesInflammation in AgingmonitamiftahNo ratings yet
- Efek Immunologi Radiasi Sinar UV MatahariDocument16 pagesEfek Immunologi Radiasi Sinar UV MataharimonitamiftahNo ratings yet
- Management of Aging SkinDocument19 pagesManagement of Aging SkinmonitamiftahNo ratings yet
- Efek Immunologi Radiasi Sinar UV MatahariDocument16 pagesEfek Immunologi Radiasi Sinar UV MataharimonitamiftahNo ratings yet
- Chapter 027Document3 pagesChapter 027barbaraNo ratings yet
- ALBCchicken Assessment 2Document6 pagesALBCchicken Assessment 2Filip Marian100% (1)
- Quick Quiz! 7-5: CHAPTER 7 Miscellaneous ProtozoaDocument17 pagesQuick Quiz! 7-5: CHAPTER 7 Miscellaneous ProtozoaTawfeeq AuqbiNo ratings yet
- Rash FeverDocument1 pageRash FeverSyima MnnNo ratings yet
- Farmacología de ItraconazolDocument5 pagesFarmacología de ItraconazolalexqaNo ratings yet
- Anatomy Physiology of The BrainDocument107 pagesAnatomy Physiology of The BrainRadha RamineniNo ratings yet
- Extraordinary Master VesselsDocument11 pagesExtraordinary Master VesselssillymindsNo ratings yet
- Exclusive Sellers of Chanson Bleu ChartreuxDocument9 pagesExclusive Sellers of Chanson Bleu ChartreuxpsykosomatikNo ratings yet
- Activity 8.2.4 Brain Anatomy Answer Key: Parietal LobeDocument5 pagesActivity 8.2.4 Brain Anatomy Answer Key: Parietal LobeAlexis CarranzaNo ratings yet
- Tài liệu học thêm 12A4 Dop: 15/3/2021 Revision (Vocabualary) I.Review 1.1 .Read the passageDocument10 pagesTài liệu học thêm 12A4 Dop: 15/3/2021 Revision (Vocabualary) I.Review 1.1 .Read the passageNguyen Thi Phuong ThaoNo ratings yet
- Solutions For Genetics Problems - Bil 151: Parental Genotypes: Male: FemaleDocument10 pagesSolutions For Genetics Problems - Bil 151: Parental Genotypes: Male: FemaleLevy Necesito0% (1)
- Usbn Bahasa Inggris Sma It Nurul Muhajirin Kotanegara Sungkai UtaraDocument5 pagesUsbn Bahasa Inggris Sma It Nurul Muhajirin Kotanegara Sungkai UtaraNuryani AhmadNo ratings yet
- Diagnosis of Mycotoxicoses in Veterinary MedicineDocument10 pagesDiagnosis of Mycotoxicoses in Veterinary MedicineDamir SuljevicNo ratings yet
- Care of Patients With Chest TubesDocument2 pagesCare of Patients With Chest Tubesaurezea100% (1)
- A Dictionary of Every Day WantsDocument571 pagesA Dictionary of Every Day Wantsdonald1976No ratings yet
- Paediatrics State Exam QuestionsDocument4 pagesPaediatrics State Exam Questionsnasibdin100% (1)
- Urine Procedure WordDocument27 pagesUrine Procedure WordYashly VargheseNo ratings yet
- 2007 Skeletal Inventory Forms (Complete)Document15 pages2007 Skeletal Inventory Forms (Complete)Sergio F Cano100% (1)
- Cva CPDocument53 pagesCva CPApol Pen100% (2)
- 1.andersen: 2. BimlerDocument6 pages1.andersen: 2. BimlerRoxana ElenaNo ratings yet
- Anal Fissure1Document3 pagesAnal Fissure1Jehanzeb ZebNo ratings yet
- BLOODDocument1 pageBLOODJuana IlaganNo ratings yet
- Wirth (2016) Core Stability in Athletes A Critical Analysis of Current PDFDocument14 pagesWirth (2016) Core Stability in Athletes A Critical Analysis of Current PDFBiology PageNo ratings yet
- Pneumocystis PneumoniaDocument5 pagesPneumocystis Pneumoniadger11No ratings yet
- Protest Petition For Establishment of KVIC National Head Quarter in Capital of India - Abhishek KadyanDocument52 pagesProtest Petition For Establishment of KVIC National Head Quarter in Capital of India - Abhishek KadyanNaresh KadyanNo ratings yet
- Electrosurgery in Periodontics PerioDocument29 pagesElectrosurgery in Periodontics PerioFourthMolar.com100% (1)
- Karya Tulis IlmiahDocument50 pagesKarya Tulis IlmiahSayyid SultanNo ratings yet