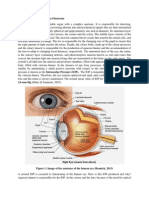
Download as pdf or txt
You might also like
- Case Presentation BronchiectasisDocument61 pagesCase Presentation BronchiectasisUzair Muhd100% (6)
- Atlas of Cardiovascular Computed Tomography - Imaging Companion To Braunwald's Heart Disease, 1E (2010) (PDF) (UnitedVRG) PDFDocument270 pagesAtlas of Cardiovascular Computed Tomography - Imaging Companion To Braunwald's Heart Disease, 1E (2010) (PDF) (UnitedVRG) PDFboghezloredanaNo ratings yet
- Reproductive System WorksheetDocument2 pagesReproductive System WorksheetEldie Ocariza100% (1)
- Functional MorphologyDocument12 pagesFunctional MorphologyNelly ElizabethNo ratings yet
- Understanding Trabecular Meshwork Physiology: A Key To The Control of Intraocular Pressure?Document5 pagesUnderstanding Trabecular Meshwork Physiology: A Key To The Control of Intraocular Pressure?RaissaNo ratings yet
- Glaucoma EssayDocument5 pagesGlaucoma Essaysujataravi6No ratings yet
- Pathogenesis of Glaucoma: Determinants of Intraocular PressureDocument17 pagesPathogenesis of Glaucoma: Determinants of Intraocular PressureAdi TriNo ratings yet
- Aqueous Humor Production 1Document7 pagesAqueous Humor Production 1yesi eka molitaNo ratings yet
- Discuss Physiology of Aqueous Formation and Its Drainage and Relate It To How Intraocular Pressure Is ControlledDocument8 pagesDiscuss Physiology of Aqueous Formation and Its Drainage and Relate It To How Intraocular Pressure Is ControlledPrince ChipunguNo ratings yet
- Jurnal ET 12Document13 pagesJurnal ET 12shendi amalia putriNo ratings yet
- Etiology and Management of EdemaDocument14 pagesEtiology and Management of EdemaFavio Alvarez De BejarNo ratings yet
- A. Primary GlaucomaDocument5 pagesA. Primary GlaucomasekiannNo ratings yet
- 2012.microinvasive Glaucoma Surgery-A Review of Schlemm's Canal-Based ProceduresDocument10 pages2012.microinvasive Glaucoma Surgery-A Review of Schlemm's Canal-Based ProceduresReza NaghibiNo ratings yet
- Understanding Trabecular Meshwork Physiology: A Key To The Control of Intraocular Pressure?Document5 pagesUnderstanding Trabecular Meshwork Physiology: A Key To The Control of Intraocular Pressure?Nail AlheloNo ratings yet
- Etiology and Management of Edema ADocument14 pagesEtiology and Management of Edema ASurco Mamani AdalidNo ratings yet
- Dr. Padmaja PhadeDocument46 pagesDr. Padmaja PhadeElaine ChongNo ratings yet
- In An: Carl KollerDocument10 pagesIn An: Carl KollerMuhammah ArslanNo ratings yet
- Osmosis FlowDocument11 pagesOsmosis FlowLaura Juliana Builes PerezNo ratings yet
- Renal PhysiologyDocument8 pagesRenal PhysiologyAlejandra Bautista OsorioNo ratings yet
- Wave Propagation and Induced Steady Streaming in Viscous Fluid Contained in A Prestressed Viscoelastic TubeDocument26 pagesWave Propagation and Induced Steady Streaming in Viscous Fluid Contained in A Prestressed Viscoelastic Tubebiswarupmandal0104No ratings yet
- Anatomy: Initiation Factors Important To Which Are Believed To Be Axial StreamDocument11 pagesAnatomy: Initiation Factors Important To Which Are Believed To Be Axial StreamogangurelNo ratings yet
- Distal Convoluted Tubules, Counter Current Mechanism, MicturitionDocument94 pagesDistal Convoluted Tubules, Counter Current Mechanism, MicturitionSarah BirechNo ratings yet
- The Electrochemical Basis of Corneal Hydration, Swelling, and TransparencyDocument18 pagesThe Electrochemical Basis of Corneal Hydration, Swelling, and TransparencyXiChengNo ratings yet
- Aqueous Humour: Anatomy & Physiology: by Tahir Muneeb - September 7, 2012 6 3502Document9 pagesAqueous Humour: Anatomy & Physiology: by Tahir Muneeb - September 7, 2012 6 3502nnurulhudaaNo ratings yet
- Careful With That Colloid, Eugene..Document36 pagesCareful With That Colloid, Eugene..Tom WoodcockNo ratings yet
- D ArteryDocument2 pagesD ArteryogangurelNo ratings yet
- Acquired Nasolacrimal Duct ObstructionDocument21 pagesAcquired Nasolacrimal Duct ObstructionLovely PoppyNo ratings yet
- 3 The Electrochemical Basis of Nerve FunctionDocument33 pages3 The Electrochemical Basis of Nerve FunctionEvets JarusNo ratings yet
- Intraocular Pressure and Aqueous Humor DynamicsDocument36 pagesIntraocular Pressure and Aqueous Humor DynamicsIntan EkarulitaNo ratings yet
- S. Faubel and J. Topf 3 Starling's LawDocument14 pagesS. Faubel and J. Topf 3 Starling's LawJoel Topf100% (5)
- Aqueous Humor Outflow:: Anterior Cavity: The Lens Divides The Anterior Cavity Into Further Two ChambersDocument20 pagesAqueous Humor Outflow:: Anterior Cavity: The Lens Divides The Anterior Cavity Into Further Two ChambersNavjot BrarNo ratings yet
- VitreousDocument37 pagesVitreousIhsanluthfi AbdillahNo ratings yet
- EpiphoraDocument17 pagesEpiphoraDhivya SekarNo ratings yet
- D Art2Document2 pagesD Art2ogangurelNo ratings yet
- Review Article: Modeling Transport and Flow Regulatory Mechanisms of The KidneyDocument19 pagesReview Article: Modeling Transport and Flow Regulatory Mechanisms of The KidneyTit fuckerNo ratings yet
- Glaucoma OverviewDocument33 pagesGlaucoma Overviewc/risaaq yuusuf ColoowNo ratings yet
- Physiology of Aqueous HumorDocument34 pagesPhysiology of Aqueous HumorCindy Seow Sieng TengNo ratings yet
- GlaucomaDocument57 pagesGlaucomaAmmad ShahidNo ratings yet
- Misdirection SyndromeDocument20 pagesMisdirection SyndromeEduardo TarazonaNo ratings yet
- CAMP Stimulates Transepithelial Short-Circuit Current andDocument11 pagesCAMP Stimulates Transepithelial Short-Circuit Current andShashi kant ChaudharyNo ratings yet
- Strasinger AUBFDocument44 pagesStrasinger AUBFangela tanteoNo ratings yet
- Anterior ChamberDocument11 pagesAnterior Chamberhanidya fazwatNo ratings yet
- Case Report Absolute GlaucomaDocument22 pagesCase Report Absolute GlaucomaDewi Permatasari100% (2)
- Wootton2001 Article AMechanisticModelOfAcutePlatelDocument9 pagesWootton2001 Article AMechanisticModelOfAcutePlatelsamik4uNo ratings yet
- Urine Concentration and DilutionDocument7 pagesUrine Concentration and DilutionHussein Ali AljamaliNo ratings yet
- A Rigorous Cake Filtration Theory: (Accepfed 5 MarchDocument12 pagesA Rigorous Cake Filtration Theory: (Accepfed 5 MarchpravkovoilaNo ratings yet
- 2015-Colocalization of Outflow Segmentation and Pores Along The Inner Wall of Schlemm's Canal. #Document46 pages2015-Colocalization of Outflow Segmentation and Pores Along The Inner Wall of Schlemm's Canal. #Reza NaghibiNo ratings yet
- The Adaptation of Altemeier's Procedure To Treat End Colostomy Prolapse: A Simple Option For Day SurgeryDocument3 pagesThe Adaptation of Altemeier's Procedure To Treat End Colostomy Prolapse: A Simple Option For Day Surgeryyohelio sibuNo ratings yet
- Cornea PhysiologyDocument76 pagesCornea Physiologydr_dev50% (2)
- Bir 1987 24606Document7 pagesBir 1987 24606Constanza Martinez ArancibiaNo ratings yet
- Gravelle 2014Document7 pagesGravelle 2014lgaliziaNo ratings yet
- Malgor 2013Document4 pagesMalgor 2013vas25No ratings yet
- Anatomy and Physiology of Cornea: Dr. Joseph JohnDocument46 pagesAnatomy and Physiology of Cornea: Dr. Joseph JohnBIBI MOHANAN100% (1)
- 2001.ageing of Schlemm's Canal in Nonglaucomatous SubjectsDocument12 pages2001.ageing of Schlemm's Canal in Nonglaucomatous SubjectsReza NaghibiNo ratings yet
- Harvade College of Science, Business and Management Studies Polt 1-12, Harvade Close, Off Obada Road Abeokuta Ogun StateDocument21 pagesHarvade College of Science, Business and Management Studies Polt 1-12, Harvade Close, Off Obada Road Abeokuta Ogun StateOluwaseun TosinNo ratings yet
- Ion Transport MechanismsDocument8 pagesIon Transport MechanismsShashi kant ChaudharyNo ratings yet
- Colloid Osmometry: Elke Rudloff, DVM, DACVECC, and Rebecca Kirby, DVM, DACVIM, DACVECCDocument7 pagesColloid Osmometry: Elke Rudloff, DVM, DACVECC, and Rebecca Kirby, DVM, DACVIM, DACVECCClayton Garcia da SilvaNo ratings yet
- 5 BD PDFDocument6 pages5 BD PDFHyakuya YuuichiroNo ratings yet
- SummaryDocument2 pagesSummaryogangurelNo ratings yet
- Fundamental Principles of Ultrafiltration PDFDocument15 pagesFundamental Principles of Ultrafiltration PDFalmutaz9879No ratings yet
- Examination of Lacrimal ApparatusDocument23 pagesExamination of Lacrimal ApparatusDr. T. BalasubramanianNo ratings yet
- Kinesiología y Sus AlcancesDocument16 pagesKinesiología y Sus AlcancesEzequiel SosaNo ratings yet
- Suprachoroidal Space InterventionsFrom EverandSuprachoroidal Space InterventionsShohista SaidkasimovaNo ratings yet
- Histology: First Semester A.Y. 2021 - 2022Document6 pagesHistology: First Semester A.Y. 2021 - 2022Fea Kristine PacquiaoNo ratings yet
- Concept II Training ManualDocument31 pagesConcept II Training Manualscribd-123No ratings yet
- Air Velocity Influences Thermoregulation and Endurance Exercise Capacity in The HeatDocument9 pagesAir Velocity Influences Thermoregulation and Endurance Exercise Capacity in The HeatMaríaNo ratings yet
- ArchRheumatol 32 21Document5 pagesArchRheumatol 32 21indriNo ratings yet
- Morphologic Evaluation of Erythrocytes: Click To Edit Master Subtitle StyleDocument41 pagesMorphologic Evaluation of Erythrocytes: Click To Edit Master Subtitle StyleRio Evonne GabayNo ratings yet
- Relapsing Polychondritis - A 2016 Update On Clinical Features, Diagnostic Tools, Treatment and Biological Drug UseDocument18 pagesRelapsing Polychondritis - A 2016 Update On Clinical Features, Diagnostic Tools, Treatment and Biological Drug UseAldiNo ratings yet
- TLSM Chapter 10 - BurnsDocument34 pagesTLSM Chapter 10 - BurnsNurul Nadia ZahariNo ratings yet
- MPSC Rajyaseva Syllabus 2023Document74 pagesMPSC Rajyaseva Syllabus 2023PravinNo ratings yet
- CARDIOPULMONARY BYPASS - CompressedDocument117 pagesCARDIOPULMONARY BYPASS - CompressedBayan TahaNo ratings yet
- Photosynthesis and Respiration WorksheetDocument2 pagesPhotosynthesis and Respiration WorksheetTyrone WaltersNo ratings yet
- Practical Proteins and Amino Acids Identification PDFDocument23 pagesPractical Proteins and Amino Acids Identification PDFFarahDeebaNo ratings yet
- Clinical ChemistryDocument7 pagesClinical Chemistrysciencetopic4No ratings yet
- Patients With Advanced Lung Cancer: Quality of Life and Perception of DyspneaDocument14 pagesPatients With Advanced Lung Cancer: Quality of Life and Perception of DyspneaSireNo ratings yet
- 7mm Frog Embryo - EMBLabDocument3 pages7mm Frog Embryo - EMBLabIvy Cruz100% (1)
- HaematologyDocument11 pagesHaematologyIkram AzmanNo ratings yet
- Chemotherapy and Renal Andhepatic Insufficiency 2008Document24 pagesChemotherapy and Renal Andhepatic Insufficiency 2008Vika RatuNo ratings yet
- J.A. Gray's Reinforcement Sensitivity Theory (RST) of PersonalityDocument18 pagesJ.A. Gray's Reinforcement Sensitivity Theory (RST) of PersonalityZoran RapaicNo ratings yet
- Chapters 7-9 - Biomolecule PresentationsDocument21 pagesChapters 7-9 - Biomolecule Presentationsapi-457044119No ratings yet
- Seeleys Essentials of Anatomy and Physiology 9th Edition Vanputte Test BankDocument38 pagesSeeleys Essentials of Anatomy and Physiology 9th Edition Vanputte Test Bankriaozgas3023100% (17)
- Pathophysiology and Etiology of Edema in AdultsDocument15 pagesPathophysiology and Etiology of Edema in AdultsJovie Anne CabangalNo ratings yet
- Cell Biology Handout - 1Document62 pagesCell Biology Handout - 1Dewi Lasimpara100% (1)
- 01 - Decision Making in Emergency MedicineDocument4 pages01 - Decision Making in Emergency MedicineSlvdrPlgrnNo ratings yet
- OTC Medication For Common Gastrointestinal ProblemsDocument3 pagesOTC Medication For Common Gastrointestinal Problemsnishanthprasad24No ratings yet
- DrugDocument6 pagesDrugRose Ann MirafloresNo ratings yet
- Digestive System ReviewDocument47 pagesDigestive System ReviewMartina RizkiNo ratings yet
- The Role of Free Radicals in Health and DiseaseDocument9 pagesThe Role of Free Radicals in Health and Diseasehumera0% (1)
- Lecture (Antibiotics) PDFDocument97 pagesLecture (Antibiotics) PDFAnonymous FMCCRgvNo ratings yet