
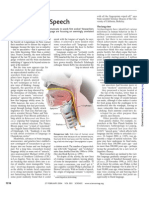
Music, Evolution and Language: Nobuo Masataka
Music, Evolution and Language: Nobuo Masataka
You might also like
- Holden Origin of Speech 2004Document4 pagesHolden Origin of Speech 2004Charlie RiveraNo ratings yet
- Nobuo Masataka (Ed.) : The Origins of Language, Unraveling Evolutionary ForcesDocument2 pagesNobuo Masataka (Ed.) : The Origins of Language, Unraveling Evolutionary ForcesAprianaRohmanNo ratings yet
- Evolutionary Musicology PDFDocument5 pagesEvolutionary Musicology PDFAnonymous OUtcQZleTQ100% (1)
- 2016 Chapter 31 Language Development PDFDocument16 pages2016 Chapter 31 Language Development PDFJulian Caro MorenoNo ratings yet
- First Page PDFDocument1 pageFirst Page PDFMichelLe LeslieNo ratings yet
- Oikkonen2014 - Genomics Approaches To Study Musical AptitudeDocument7 pagesOikkonen2014 - Genomics Approaches To Study Musical AptitudeLiliane OliveiraNo ratings yet
- The Biology and Evolution of Music (43 PG)Document43 pagesThe Biology and Evolution of Music (43 PG)ricardosmith17No ratings yet
- Interactiv Bioacoustic Playback As A Tool For Detecting and Exploring Nonhuman Intelligence "Conversing" With An Alaskan Humpback WhaleDocument21 pagesInteractiv Bioacoustic Playback As A Tool For Detecting and Exploring Nonhuman Intelligence "Conversing" With An Alaskan Humpback WhaleIsabelle Campello da MataNo ratings yet
- VARC1000 - CAD 41 To 47Document63 pagesVARC1000 - CAD 41 To 47Keshav BahetyNo ratings yet
- Music and Dance in MammalsDocument4 pagesMusic and Dance in Mammalspeteycosta100% (1)
- Doupe, A. J., & Kuhl, P. K. (1999) - Birdsong and Human Speech - Common Themes and Mechanisms. Annual Review of Neuroscience, 22 (1), 567-631.Document66 pagesDoupe, A. J., & Kuhl, P. K. (1999) - Birdsong and Human Speech - Common Themes and Mechanisms. Annual Review of Neuroscience, 22 (1), 567-631.goni56509No ratings yet
- The Biology and Evolution of Music: A Comparative PerspectiveDocument43 pagesThe Biology and Evolution of Music: A Comparative PerspectiveMakai Péter KristófNo ratings yet
- Vocalizations of Infants - 17nov10Document30 pagesVocalizations of Infants - 17nov10prof.joao.reigadoNo ratings yet
- 2011 Evolutionof Phonology Lindblom Camb Encyclo LG SciDocument4 pages2011 Evolutionof Phonology Lindblom Camb Encyclo LG SciaifasimahetsafiatehamiNo ratings yet
- Is Language Acquisition Learned or Inbuilt? Exploration of Nature Versus Nurture DebateDocument10 pagesIs Language Acquisition Learned or Inbuilt? Exploration of Nature Versus Nurture DebateHediliza TomeldenNo ratings yet
- Sharing Neural Networks: Early Music Training Enhances Language DevelopmentDocument13 pagesSharing Neural Networks: Early Music Training Enhances Language DevelopmentJessica ComstockNo ratings yet
- DeveThe Development of Speaking and Singing in Infants May Playa Role in Genomics and Dementia in Humans Page 4Document1 pageDeveThe Development of Speaking and Singing in Infants May Playa Role in Genomics and Dementia in Humans Page 4yoxoxec954No ratings yet
- Perspectives: An Open Introduction To Cultural Anthropology: Second EditionDocument28 pagesPerspectives: An Open Introduction To Cultural Anthropology: Second EditionAnanya samuelNo ratings yet
- Birdsong and Human SpeechDocument66 pagesBirdsong and Human SpeechJennyLomaxNo ratings yet
- Vocalizations of Infants (9-11 Months Old) in Response To Musical and Linguistic StimuliDocument16 pagesVocalizations of Infants (9-11 Months Old) in Response To Musical and Linguistic StimuliLaura F MerinoNo ratings yet
- Case Study On The Origins of Language Particularly On The Natural Sound SourceDocument2 pagesCase Study On The Origins of Language Particularly On The Natural Sound Sourceロレンソ セブゲロ カジェラNo ratings yet
- Jackendoff How Did Language BeginDocument4 pagesJackendoff How Did Language BeginLUCÍA MACARENA CATALINA GONZÁLEZ-LLANOSNo ratings yet
- Zuberbu - 2005 - The Phylogenetic Roots of Language Evidence From Primate Communication and CognitionDocument5 pagesZuberbu - 2005 - The Phylogenetic Roots of Language Evidence From Primate Communication and CognitionJohnNo ratings yet
- Sign Language Versus Spoken Language: Stokoe, William CDocument20 pagesSign Language Versus Spoken Language: Stokoe, William CCrisNo ratings yet
- Nature Versus Nurture in Language AcquisitionDocument2 pagesNature Versus Nurture in Language AcquisitionnanaamoNo ratings yet
- Graham F. Welch and Costanza Preti. Singing As Inter - and Intra-Personal Communication.Document28 pagesGraham F. Welch and Costanza Preti. Singing As Inter - and Intra-Personal Communication.Юрий СемёновNo ratings yet
- Why Do We Like To Dance and Sing 2005Document16 pagesWhy Do We Like To Dance and Sing 2005muzicoterapie1092No ratings yet
- Everyday Music InfancyDocument25 pagesEveryday Music InfancySistemassocialesNo ratings yet
- A New View of Language AcquisitionDocument8 pagesA New View of Language AcquisitioncarlalopezbiologaNo ratings yet
- A34090 - Phạm Quang Vinh - CĐTN NN&VHDocument15 pagesA34090 - Phạm Quang Vinh - CĐTN NN&VHTH Lý Thái Tổ Nguyễn Thị HườngNo ratings yet
- International Encyclopedia of The Social & Behavioral - Croft, William - Pages 364-369, 2015 - ElsevDocument6 pagesInternational Encyclopedia of The Social & Behavioral - Croft, William - Pages 364-369, 2015 - ElsevAhmad MuliaNo ratings yet
- 11.kirschner Tomasello 2010 Joint Music Making Promotes Prosocial Behavior EHBDocument11 pages11.kirschner Tomasello 2010 Joint Music Making Promotes Prosocial Behavior EHBzainabNo ratings yet
- FLAandmelodysingingDocument8 pagesFLAandmelodysingingFranNo ratings yet
- Process Worksheet Origins of LanguageDocument3 pagesProcess Worksheet Origins of Languagejuhanisahjamiel6No ratings yet
- Origins of Human LanguageDocument31 pagesOrigins of Human LanguageZubair AliNo ratings yet
- Chapter 1Document4 pagesChapter 1Nikoleta RudnikNo ratings yet
- Unit 1 Evolution of Language: StructureDocument231 pagesUnit 1 Evolution of Language: StructureRochi Gadi AddaNo ratings yet
- Unit 1-Evolution of LanguageDocument12 pagesUnit 1-Evolution of LanguageShahana ThasinNo ratings yet
- Nilayda Karakılıç - The Musical, Social Interaction and Physical Adaptation Source of LanguageDocument35 pagesNilayda Karakılıç - The Musical, Social Interaction and Physical Adaptation Source of LanguageNilayda KarakılıçNo ratings yet
- NativismDocument4 pagesNativismKeiros FuegoNo ratings yet
- The Biology and Evolution of Music: A Comparative PerspectiveDocument43 pagesThe Biology and Evolution of Music: A Comparative PerspectiveHoja de ChopoNo ratings yet
- (Emerson) Origin of LanguageDocument14 pages(Emerson) Origin of LanguageSami ktkNo ratings yet
- Musical Predispositions in InfancyDocument17 pagesMusical Predispositions in InfancydanielabanderasNo ratings yet
- Mothers and Infants and Communicative MusicalityDocument30 pagesMothers and Infants and Communicative MusicalityJoão Pedro ReigadoNo ratings yet
- The Origins of LanguageDocument7 pagesThe Origins of Languagefur94fadNo ratings yet
- Unige 102028 Attachment01Document15 pagesUnige 102028 Attachment01supiboboNo ratings yet
- Beyaz Kahverengi Minimal Kahvenin Tarihi SunumDocument29 pagesBeyaz Kahverengi Minimal Kahvenin Tarihi SunumNilayda KarakılıçNo ratings yet
- Development of The English LanguageDocument2 pagesDevelopment of The English LanguageJane ButalidNo ratings yet
- Chapter 4Document34 pagesChapter 4Teerach SongwatanaNo ratings yet
- Fondaction of LanguageDocument11 pagesFondaction of LanguagecarmenNo ratings yet
- Language Origins and DiversificationDocument2 pagesLanguage Origins and DiversificationgiustamarcoNo ratings yet
- MeaningS of SoundDocument9 pagesMeaningS of SoundRobert ReigleNo ratings yet
- Differences in Acoustic Features of Vocalizations Killer Whales Dolphins COMUNICACIÓN CETÁCEOS Musser 14 PDFDocument13 pagesDifferences in Acoustic Features of Vocalizations Killer Whales Dolphins COMUNICACIÓN CETÁCEOS Musser 14 PDFGogelioNo ratings yet
- Música y Lenguaje ComparaciónDocument23 pagesMúsica y Lenguaje Comparaciónmartha diazNo ratings yet
- The Origins of Language: Human Responses To SoundsDocument2 pagesThe Origins of Language: Human Responses To SoundsAsad MehmoodNo ratings yet
- Music-Where Cognition and Emotion MeetDocument7 pagesMusic-Where Cognition and Emotion MeetLílian OLiveira SalesNo ratings yet
- HBM 21173 PDFDocument16 pagesHBM 21173 PDFJovy EsquivelNo ratings yet
- Manpreet Kaur (23101405)Document15 pagesManpreet Kaur (23101405)preetbhangu074No ratings yet
- Introduction: Integrating Genetic and Cultural Evolutionary Approaches To LanguageDocument12 pagesIntroduction: Integrating Genetic and Cultural Evolutionary Approaches To LanguageVíctor Julián VallejoNo ratings yet
- Types of ShoringDocument9 pagesTypes of ShoringAzil14100% (10)
- The Advantages of Fiedler's Contingency ModelDocument10 pagesThe Advantages of Fiedler's Contingency ModelAnwar LaskarNo ratings yet
- C.S. Sharma and Kamlesh Purohit - Theory of Mechanisms and Machines-PHI Learning Private Limited (2006)Document723 pagesC.S. Sharma and Kamlesh Purohit - Theory of Mechanisms and Machines-PHI Learning Private Limited (2006)tp coephNo ratings yet
- CA Level 1Document43 pagesCA Level 1Cikya ComelNo ratings yet
- SR Designworks: Head OfficeDocument15 pagesSR Designworks: Head Officeihameed4100% (1)
- StudyGuide CosmonautDocument25 pagesStudyGuide CosmonautAzuWillDieNo ratings yet
- 3VA22255HL420AA0 Datasheet enDocument7 pages3VA22255HL420AA0 Datasheet enBensonNo ratings yet
- Rudder - Steering Gear Speed RulesDNVGL-RU-SHIP-Pt4Ch10 19Document1 pageRudder - Steering Gear Speed RulesDNVGL-RU-SHIP-Pt4Ch10 19Tolias EgwNo ratings yet
- Installation and Operation Manual: VRV IV+ Heat Pump For High Ambient TemperaturesDocument44 pagesInstallation and Operation Manual: VRV IV+ Heat Pump For High Ambient Temperaturesahmadgce04No ratings yet
- Community Involvement Output The Importance of Community Development ProjectsDocument7 pagesCommunity Involvement Output The Importance of Community Development ProjectsMark Angelo NasNo ratings yet
- Floating Boat Library-Mini Project-1Document37 pagesFloating Boat Library-Mini Project-1parmayadav56789No ratings yet
- Lorentz Factor Formula - Google SearchDocument1 pageLorentz Factor Formula - Google Searchnppptry4kmNo ratings yet
- ISA75X75X8 X 765: Asset Development Plan 1 (ADP-1)Document1 pageISA75X75X8 X 765: Asset Development Plan 1 (ADP-1)Tamil Arasu SNo ratings yet
- Adu Gloria WorkDocument68 pagesAdu Gloria WorkAiman UsmanNo ratings yet
- Syllabus For Applied ElectronicsDocument28 pagesSyllabus For Applied ElectronicsvinayakbondNo ratings yet
- 2.High-Confidence Behavior at Work The Exec SkillsDocument27 pages2.High-Confidence Behavior at Work The Exec SkillsRamiro BernalesNo ratings yet
- Activity Sheets: Quarter 3 - MELC 2Document9 pagesActivity Sheets: Quarter 3 - MELC 2Alexa HullezaNo ratings yet
- Chapter 1: Scientific Understanding of Behavior: Learning ObjectivesDocument13 pagesChapter 1: Scientific Understanding of Behavior: Learning Objectiveshallo BroNo ratings yet
- My First 10 Years ProjectDocument2 pagesMy First 10 Years ProjectahdyalNo ratings yet
- MC Practicals 2Document12 pagesMC Practicals 2Adi AdnanNo ratings yet
- ADT Service ManualDocument152 pagesADT Service ManualZakhele MpofuNo ratings yet
- Journal of Art, Architecture and Built Environment (JAABE)Document15 pagesJournal of Art, Architecture and Built Environment (JAABE)UMT JournalsNo ratings yet
- Elizabeth Winifred Brewster,: CanadianDocument4 pagesElizabeth Winifred Brewster,: CanadianVinnoDarmawanNo ratings yet
- SEAS RSRC 57bfe799620e0Document4 pagesSEAS RSRC 57bfe799620e0WunNaNo ratings yet
- Unit 5 - Event Driven ProgrammingDocument22 pagesUnit 5 - Event Driven ProgrammingdharanyaNo ratings yet
- Gear Agma IIDocument49 pagesGear Agma IInathNo ratings yet
- SYNTHESIS Filipino 1Document6 pagesSYNTHESIS Filipino 1DhanNo ratings yet
- Complete Retro Kenner Action Figure Checklist - ReAction, Warpo, Amok Time, Zica ToysDocument6 pagesComplete Retro Kenner Action Figure Checklist - ReAction, Warpo, Amok Time, Zica ToysJadBeanNo ratings yet
- Author's Purpose Mini PassagesDocument5 pagesAuthor's Purpose Mini Passages18118No ratings yet
- Nynas Transformer Oil - Nytro 10GBN: Naphthenics Product Data SheetDocument1 pageNynas Transformer Oil - Nytro 10GBN: Naphthenics Product Data SheetAnonymous S29FwnFNo ratings yet
























































































Download as pdf or txt
You might also like
- Holden Origin of Speech 2004Document4 pagesHolden Origin of Speech 2004Charlie RiveraNo ratings yet
- Nobuo Masataka (Ed.) : The Origins of Language, Unraveling Evolutionary ForcesDocument2 pagesNobuo Masataka (Ed.) : The Origins of Language, Unraveling Evolutionary ForcesAprianaRohmanNo ratings yet
- Evolutionary Musicology PDFDocument5 pagesEvolutionary Musicology PDFAnonymous OUtcQZleTQ100% (1)
- 2016 Chapter 31 Language Development PDFDocument16 pages2016 Chapter 31 Language Development PDFJulian Caro MorenoNo ratings yet
- First Page PDFDocument1 pageFirst Page PDFMichelLe LeslieNo ratings yet
- Oikkonen2014 - Genomics Approaches To Study Musical AptitudeDocument7 pagesOikkonen2014 - Genomics Approaches To Study Musical AptitudeLiliane OliveiraNo ratings yet
- The Biology and Evolution of Music (43 PG)Document43 pagesThe Biology and Evolution of Music (43 PG)ricardosmith17No ratings yet
- Interactiv Bioacoustic Playback As A Tool For Detecting and Exploring Nonhuman Intelligence "Conversing" With An Alaskan Humpback WhaleDocument21 pagesInteractiv Bioacoustic Playback As A Tool For Detecting and Exploring Nonhuman Intelligence "Conversing" With An Alaskan Humpback WhaleIsabelle Campello da MataNo ratings yet
- VARC1000 - CAD 41 To 47Document63 pagesVARC1000 - CAD 41 To 47Keshav BahetyNo ratings yet
- Music and Dance in MammalsDocument4 pagesMusic and Dance in Mammalspeteycosta100% (1)
- Doupe, A. J., & Kuhl, P. K. (1999) - Birdsong and Human Speech - Common Themes and Mechanisms. Annual Review of Neuroscience, 22 (1), 567-631.Document66 pagesDoupe, A. J., & Kuhl, P. K. (1999) - Birdsong and Human Speech - Common Themes and Mechanisms. Annual Review of Neuroscience, 22 (1), 567-631.goni56509No ratings yet
- The Biology and Evolution of Music: A Comparative PerspectiveDocument43 pagesThe Biology and Evolution of Music: A Comparative PerspectiveMakai Péter KristófNo ratings yet
- Vocalizations of Infants - 17nov10Document30 pagesVocalizations of Infants - 17nov10prof.joao.reigadoNo ratings yet
- 2011 Evolutionof Phonology Lindblom Camb Encyclo LG SciDocument4 pages2011 Evolutionof Phonology Lindblom Camb Encyclo LG SciaifasimahetsafiatehamiNo ratings yet
- Is Language Acquisition Learned or Inbuilt? Exploration of Nature Versus Nurture DebateDocument10 pagesIs Language Acquisition Learned or Inbuilt? Exploration of Nature Versus Nurture DebateHediliza TomeldenNo ratings yet
- Sharing Neural Networks: Early Music Training Enhances Language DevelopmentDocument13 pagesSharing Neural Networks: Early Music Training Enhances Language DevelopmentJessica ComstockNo ratings yet
- DeveThe Development of Speaking and Singing in Infants May Playa Role in Genomics and Dementia in Humans Page 4Document1 pageDeveThe Development of Speaking and Singing in Infants May Playa Role in Genomics and Dementia in Humans Page 4yoxoxec954No ratings yet
- Perspectives: An Open Introduction To Cultural Anthropology: Second EditionDocument28 pagesPerspectives: An Open Introduction To Cultural Anthropology: Second EditionAnanya samuelNo ratings yet
- Birdsong and Human SpeechDocument66 pagesBirdsong and Human SpeechJennyLomaxNo ratings yet
- Vocalizations of Infants (9-11 Months Old) in Response To Musical and Linguistic StimuliDocument16 pagesVocalizations of Infants (9-11 Months Old) in Response To Musical and Linguistic StimuliLaura F MerinoNo ratings yet
- Case Study On The Origins of Language Particularly On The Natural Sound SourceDocument2 pagesCase Study On The Origins of Language Particularly On The Natural Sound Sourceロレンソ セブゲロ カジェラNo ratings yet
- Jackendoff How Did Language BeginDocument4 pagesJackendoff How Did Language BeginLUCÍA MACARENA CATALINA GONZÁLEZ-LLANOSNo ratings yet
- Zuberbu - 2005 - The Phylogenetic Roots of Language Evidence From Primate Communication and CognitionDocument5 pagesZuberbu - 2005 - The Phylogenetic Roots of Language Evidence From Primate Communication and CognitionJohnNo ratings yet
- Sign Language Versus Spoken Language: Stokoe, William CDocument20 pagesSign Language Versus Spoken Language: Stokoe, William CCrisNo ratings yet
- Nature Versus Nurture in Language AcquisitionDocument2 pagesNature Versus Nurture in Language AcquisitionnanaamoNo ratings yet
- Graham F. Welch and Costanza Preti. Singing As Inter - and Intra-Personal Communication.Document28 pagesGraham F. Welch and Costanza Preti. Singing As Inter - and Intra-Personal Communication.Юрий СемёновNo ratings yet
- Why Do We Like To Dance and Sing 2005Document16 pagesWhy Do We Like To Dance and Sing 2005muzicoterapie1092No ratings yet
- Everyday Music InfancyDocument25 pagesEveryday Music InfancySistemassocialesNo ratings yet
- A New View of Language AcquisitionDocument8 pagesA New View of Language AcquisitioncarlalopezbiologaNo ratings yet
- A34090 - Phạm Quang Vinh - CĐTN NN&VHDocument15 pagesA34090 - Phạm Quang Vinh - CĐTN NN&VHTH Lý Thái Tổ Nguyễn Thị HườngNo ratings yet
- International Encyclopedia of The Social & Behavioral - Croft, William - Pages 364-369, 2015 - ElsevDocument6 pagesInternational Encyclopedia of The Social & Behavioral - Croft, William - Pages 364-369, 2015 - ElsevAhmad MuliaNo ratings yet
- 11.kirschner Tomasello 2010 Joint Music Making Promotes Prosocial Behavior EHBDocument11 pages11.kirschner Tomasello 2010 Joint Music Making Promotes Prosocial Behavior EHBzainabNo ratings yet
- FLAandmelodysingingDocument8 pagesFLAandmelodysingingFranNo ratings yet
- Process Worksheet Origins of LanguageDocument3 pagesProcess Worksheet Origins of Languagejuhanisahjamiel6No ratings yet
- Origins of Human LanguageDocument31 pagesOrigins of Human LanguageZubair AliNo ratings yet
- Chapter 1Document4 pagesChapter 1Nikoleta RudnikNo ratings yet
- Unit 1 Evolution of Language: StructureDocument231 pagesUnit 1 Evolution of Language: StructureRochi Gadi AddaNo ratings yet
- Unit 1-Evolution of LanguageDocument12 pagesUnit 1-Evolution of LanguageShahana ThasinNo ratings yet
- Nilayda Karakılıç - The Musical, Social Interaction and Physical Adaptation Source of LanguageDocument35 pagesNilayda Karakılıç - The Musical, Social Interaction and Physical Adaptation Source of LanguageNilayda KarakılıçNo ratings yet
- NativismDocument4 pagesNativismKeiros FuegoNo ratings yet
- The Biology and Evolution of Music: A Comparative PerspectiveDocument43 pagesThe Biology and Evolution of Music: A Comparative PerspectiveHoja de ChopoNo ratings yet
- (Emerson) Origin of LanguageDocument14 pages(Emerson) Origin of LanguageSami ktkNo ratings yet
- Musical Predispositions in InfancyDocument17 pagesMusical Predispositions in InfancydanielabanderasNo ratings yet
- Mothers and Infants and Communicative MusicalityDocument30 pagesMothers and Infants and Communicative MusicalityJoão Pedro ReigadoNo ratings yet
- The Origins of LanguageDocument7 pagesThe Origins of Languagefur94fadNo ratings yet
- Unige 102028 Attachment01Document15 pagesUnige 102028 Attachment01supiboboNo ratings yet
- Beyaz Kahverengi Minimal Kahvenin Tarihi SunumDocument29 pagesBeyaz Kahverengi Minimal Kahvenin Tarihi SunumNilayda KarakılıçNo ratings yet
- Development of The English LanguageDocument2 pagesDevelopment of The English LanguageJane ButalidNo ratings yet
- Chapter 4Document34 pagesChapter 4Teerach SongwatanaNo ratings yet
- Fondaction of LanguageDocument11 pagesFondaction of LanguagecarmenNo ratings yet
- Language Origins and DiversificationDocument2 pagesLanguage Origins and DiversificationgiustamarcoNo ratings yet
- MeaningS of SoundDocument9 pagesMeaningS of SoundRobert ReigleNo ratings yet
- Differences in Acoustic Features of Vocalizations Killer Whales Dolphins COMUNICACIÓN CETÁCEOS Musser 14 PDFDocument13 pagesDifferences in Acoustic Features of Vocalizations Killer Whales Dolphins COMUNICACIÓN CETÁCEOS Musser 14 PDFGogelioNo ratings yet
- Música y Lenguaje ComparaciónDocument23 pagesMúsica y Lenguaje Comparaciónmartha diazNo ratings yet
- The Origins of Language: Human Responses To SoundsDocument2 pagesThe Origins of Language: Human Responses To SoundsAsad MehmoodNo ratings yet
- Music-Where Cognition and Emotion MeetDocument7 pagesMusic-Where Cognition and Emotion MeetLílian OLiveira SalesNo ratings yet
- HBM 21173 PDFDocument16 pagesHBM 21173 PDFJovy EsquivelNo ratings yet
- Manpreet Kaur (23101405)Document15 pagesManpreet Kaur (23101405)preetbhangu074No ratings yet
- Introduction: Integrating Genetic and Cultural Evolutionary Approaches To LanguageDocument12 pagesIntroduction: Integrating Genetic and Cultural Evolutionary Approaches To LanguageVíctor Julián VallejoNo ratings yet
- Types of ShoringDocument9 pagesTypes of ShoringAzil14100% (10)
- The Advantages of Fiedler's Contingency ModelDocument10 pagesThe Advantages of Fiedler's Contingency ModelAnwar LaskarNo ratings yet
- C.S. Sharma and Kamlesh Purohit - Theory of Mechanisms and Machines-PHI Learning Private Limited (2006)Document723 pagesC.S. Sharma and Kamlesh Purohit - Theory of Mechanisms and Machines-PHI Learning Private Limited (2006)tp coephNo ratings yet
- CA Level 1Document43 pagesCA Level 1Cikya ComelNo ratings yet
- SR Designworks: Head OfficeDocument15 pagesSR Designworks: Head Officeihameed4100% (1)
- StudyGuide CosmonautDocument25 pagesStudyGuide CosmonautAzuWillDieNo ratings yet
- 3VA22255HL420AA0 Datasheet enDocument7 pages3VA22255HL420AA0 Datasheet enBensonNo ratings yet
- Rudder - Steering Gear Speed RulesDNVGL-RU-SHIP-Pt4Ch10 19Document1 pageRudder - Steering Gear Speed RulesDNVGL-RU-SHIP-Pt4Ch10 19Tolias EgwNo ratings yet
- Installation and Operation Manual: VRV IV+ Heat Pump For High Ambient TemperaturesDocument44 pagesInstallation and Operation Manual: VRV IV+ Heat Pump For High Ambient Temperaturesahmadgce04No ratings yet
- Community Involvement Output The Importance of Community Development ProjectsDocument7 pagesCommunity Involvement Output The Importance of Community Development ProjectsMark Angelo NasNo ratings yet
- Floating Boat Library-Mini Project-1Document37 pagesFloating Boat Library-Mini Project-1parmayadav56789No ratings yet
- Lorentz Factor Formula - Google SearchDocument1 pageLorentz Factor Formula - Google Searchnppptry4kmNo ratings yet
- ISA75X75X8 X 765: Asset Development Plan 1 (ADP-1)Document1 pageISA75X75X8 X 765: Asset Development Plan 1 (ADP-1)Tamil Arasu SNo ratings yet
- Adu Gloria WorkDocument68 pagesAdu Gloria WorkAiman UsmanNo ratings yet
- Syllabus For Applied ElectronicsDocument28 pagesSyllabus For Applied ElectronicsvinayakbondNo ratings yet
- 2.High-Confidence Behavior at Work The Exec SkillsDocument27 pages2.High-Confidence Behavior at Work The Exec SkillsRamiro BernalesNo ratings yet
- Activity Sheets: Quarter 3 - MELC 2Document9 pagesActivity Sheets: Quarter 3 - MELC 2Alexa HullezaNo ratings yet
- Chapter 1: Scientific Understanding of Behavior: Learning ObjectivesDocument13 pagesChapter 1: Scientific Understanding of Behavior: Learning Objectiveshallo BroNo ratings yet
- My First 10 Years ProjectDocument2 pagesMy First 10 Years ProjectahdyalNo ratings yet
- MC Practicals 2Document12 pagesMC Practicals 2Adi AdnanNo ratings yet
- ADT Service ManualDocument152 pagesADT Service ManualZakhele MpofuNo ratings yet
- Journal of Art, Architecture and Built Environment (JAABE)Document15 pagesJournal of Art, Architecture and Built Environment (JAABE)UMT JournalsNo ratings yet
- Elizabeth Winifred Brewster,: CanadianDocument4 pagesElizabeth Winifred Brewster,: CanadianVinnoDarmawanNo ratings yet
- SEAS RSRC 57bfe799620e0Document4 pagesSEAS RSRC 57bfe799620e0WunNaNo ratings yet
- Unit 5 - Event Driven ProgrammingDocument22 pagesUnit 5 - Event Driven ProgrammingdharanyaNo ratings yet
- Gear Agma IIDocument49 pagesGear Agma IInathNo ratings yet
- SYNTHESIS Filipino 1Document6 pagesSYNTHESIS Filipino 1DhanNo ratings yet
- Complete Retro Kenner Action Figure Checklist - ReAction, Warpo, Amok Time, Zica ToysDocument6 pagesComplete Retro Kenner Action Figure Checklist - ReAction, Warpo, Amok Time, Zica ToysJadBeanNo ratings yet
- Author's Purpose Mini PassagesDocument5 pagesAuthor's Purpose Mini Passages18118No ratings yet
- Nynas Transformer Oil - Nytro 10GBN: Naphthenics Product Data SheetDocument1 pageNynas Transformer Oil - Nytro 10GBN: Naphthenics Product Data SheetAnonymous S29FwnFNo ratings yet