Download as pdf or txt
You might also like
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5821)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1093)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (852)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (590)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (898)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (540)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (349)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (822)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (122)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (403)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (74)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- Physics Annual PlanDocument60 pagesPhysics Annual Plan112233445566778899 998877665544332211100% (2)
- Cold Call Scripts - Coaching SessionDocument3 pagesCold Call Scripts - Coaching SessionDianeNo ratings yet
- Manual of YFD300 Ultrasonic Flaw DetectorDocument72 pagesManual of YFD300 Ultrasonic Flaw DetectorHenry CruzNo ratings yet
- O-Level Commerce PDFDocument8 pagesO-Level Commerce PDFClinton ChikengezhaNo ratings yet
- Krystexxa FDADocument314 pagesKrystexxa FDAVenkata Suryanarayana GorleNo ratings yet
- Example Exclusive Patent LicenseDocument13 pagesExample Exclusive Patent LicenseSam Han100% (1)
- Ensemble LearningDocument7 pagesEnsemble LearningJavier Garcia RajoyNo ratings yet
- Dee Deng's PresentationDocument31 pagesDee Deng's PresentationИгорь МореходовNo ratings yet
- Lexi BDocument62 pagesLexi BPetruss RonyyNo ratings yet
- Null 13Document4 pagesNull 13oreeditsemasilo85No ratings yet
- Arts Activity Sheet: Quarter 2 - Week 1-2Document8 pagesArts Activity Sheet: Quarter 2 - Week 1-2Charrie Mae MalloNo ratings yet
- Backplane Designer's Guide 4Document11 pagesBackplane Designer's Guide 4terryschNo ratings yet
- Ignition Characteristics of Cracked JP-7 FuelDocument4 pagesIgnition Characteristics of Cracked JP-7 Fuelfahrul3xNo ratings yet
- Core Java InterviewDocument251 pagesCore Java InterviewLalitNo ratings yet
- Grasso: Ammonia Liquid Chiller Series FX PP, LP, VP 200 - 6000 KWDocument65 pagesGrasso: Ammonia Liquid Chiller Series FX PP, LP, VP 200 - 6000 KWManuel MartinezNo ratings yet
- Book 14 PDFDocument242 pagesBook 14 PDFAndres Perez71% (7)
- Vastu-Truths and UntruthsDocument5 pagesVastu-Truths and Untruthsbas daNo ratings yet
- Math NCF 268-293Document26 pagesMath NCF 268-293natureyt4No ratings yet
- BBRIP - Monitoring ToolDocument2 pagesBBRIP - Monitoring ToolGeraldine N. VitoNo ratings yet
- Math ProblemsDocument3 pagesMath Problemsaibajo2004No ratings yet
- Akashic Brotherhood 4 PrintDocument33 pagesAkashic Brotherhood 4 Printmdamascus75643No ratings yet
- Parker Valvula Direcional D3W PDFDocument20 pagesParker Valvula Direcional D3W PDFChagas OliveiraNo ratings yet
- User Manual IvmsDocument223 pagesUser Manual IvmsGerardo AltobelliNo ratings yet
- Basic Customer Service Principles-ImportantDocument39 pagesBasic Customer Service Principles-Importantzoltan2014100% (6)
- 600 ngữ pháp chọn lọc luyện thi THPT Quốc GiaDocument32 pages600 ngữ pháp chọn lọc luyện thi THPT Quốc GiaNguyen Chi Cuong100% (1)
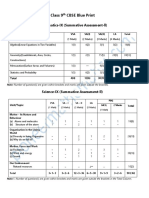
- Class 9th CBSE Blue PrintDocument2 pagesClass 9th CBSE Blue PrintYash BawiskarNo ratings yet
- Comparatives and Superlatives Interactive PracticeDocument5 pagesComparatives and Superlatives Interactive Practiceyenifer sanchezNo ratings yet
- Patient NameDocument3 pagesPatient NameSmith91No ratings yet
- Ip 15Document42 pagesIp 15owenh796100% (4)
- Muhammad Farid Ubaidillah - Youth's EndeavorDocument4 pagesMuhammad Farid Ubaidillah - Youth's EndeavorMuhammad farid ubaidillah UB 2022No ratings yet