
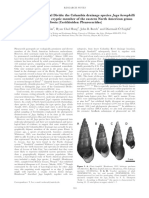
SZ Alan Ski 2003
SZ Alan Ski 2003
You might also like
- Colorado Flora: Eastern Slope, Fourth Edition A Field Guide to the Vascular PlantsFrom EverandColorado Flora: Eastern Slope, Fourth Edition A Field Guide to the Vascular PlantsRating: 3.5 out of 5 stars3.5/5 (2)
- Power of GravityDocument71 pagesPower of GravityJose RodriguezNo ratings yet
- Adjuvants For Agrichemicals PDFDocument764 pagesAdjuvants For Agrichemicals PDFAlvaro ProañoNo ratings yet
- Ua School of Journalism Professional Project Proposal Guidelines For The Master'S Degree ProgramDocument8 pagesUa School of Journalism Professional Project Proposal Guidelines For The Master'S Degree ProgramChaii Madlangsakay TolentinoNo ratings yet
- Project Report On LakmeDocument53 pagesProject Report On LakmeOmita Johar69% (13)
- South Florida Vegetable Pest and Disease Hotline For January 3, 2018Document24 pagesSouth Florida Vegetable Pest and Disease Hotline For January 3, 2018Gene McAvoyNo ratings yet
- Insecticide Resistance of Diamondback Moth (Lepidoptera: Plutellidae) in North AmericaDocument9 pagesInsecticide Resistance of Diamondback Moth (Lepidoptera: Plutellidae) in North AmericaFlorencia GorordoNo ratings yet
- 314Document4 pages314Carlos MeirellesNo ratings yet
- Castilleja: A Publication of The Wyoming Native Plant SocietyDocument12 pagesCastilleja: A Publication of The Wyoming Native Plant SocietyWyoming Native Plant SocietyNo ratings yet
- Trout Production - United States Department of AgricultureDocument15 pagesTrout Production - United States Department of AgriculturehighmountainbearNo ratings yet
- Arnett Thomas Skelley Frank 2002Document878 pagesArnett Thomas Skelley Frank 2002raitomi04No ratings yet
- YoungEtAl ChenopodDocument17 pagesYoungEtAl ChenopodCHrisNo ratings yet
- Castilleja: A Publication of The Wyoming Native Plant SocietyDocument12 pagesCastilleja: A Publication of The Wyoming Native Plant SocietyWyoming Native Plant SocietyNo ratings yet
- Fishes of North America Endangered, Threatened, or of Special Concern: 1979Document16 pagesFishes of North America Endangered, Threatened, or of Special Concern: 1979guillermo lopezNo ratings yet
- Cut Flowers and FoliagesFrom EverandCut Flowers and FoliagesJames E. FaustNo ratings yet
- Colorado Flora: Western Slope, Fourth Edition A Field Guide to the Vascular PlantsFrom EverandColorado Flora: Western Slope, Fourth Edition A Field Guide to the Vascular PlantsNo ratings yet
- Castilleja: A Publication of The Wyoming Native Plant SocietyDocument10 pagesCastilleja: A Publication of The Wyoming Native Plant SocietyWyoming Native Plant SocietyNo ratings yet
- CDTX PatentDocument364 pagesCDTX PatentCharles GrossNo ratings yet
- American Beetles, v.2 (Arnett Et Al., 2002)Document881 pagesAmerican Beetles, v.2 (Arnett Et Al., 2002)Paco Ramirez100% (1)
- Scholtz Decapod 2009 PDFDocument22 pagesScholtz Decapod 2009 PDFJaqen H'gharNo ratings yet
- Bringraj Eclipta Alba ECPRDocument3 pagesBringraj Eclipta Alba ECPRRajesh KumarNo ratings yet
- Giant Water Bug in AsiaDocument11 pagesGiant Water Bug in AsiaDũng Hoàng TrầnNo ratings yet
- Special Sego Lily Internet Offer For All Members!: NOV/DEC 2002 NOV/DEC 2002Document8 pagesSpecial Sego Lily Internet Offer For All Members!: NOV/DEC 2002 NOV/DEC 2002Friends of Utah Native Plant SocietyNo ratings yet
- Sove 1993, Vol 18, No 2 PDFDocument124 pagesSove 1993, Vol 18, No 2 PDFanggit prihatnoloNo ratings yet
- Jashs Article p554Document4 pagesJashs Article p554raraNo ratings yet
- Pdis-93-6-0584.pdf PatooDocument9 pagesPdis-93-6-0584.pdf PatooJose Miguel Soto HerediaNo ratings yet
- Gao - Et.al-2023-The Generation of The First Chromosome-Level de Novo Genome Assembly and Development and validation-50KSNParray-SalmonDocument14 pagesGao - Et.al-2023-The Generation of The First Chromosome-Level de Novo Genome Assembly and Development and validation-50KSNParray-SalmonRama BangeraNo ratings yet
- Notice of Release DS5-67 (Soybean)Document3 pagesNotice of Release DS5-67 (Soybean)Rodrigo Vazquez ValdezNo ratings yet
- Botany Labels Final FinalDocument11 pagesBotany Labels Final FinalAlexis MoffNo ratings yet
- What'S Inside: Julie PondDocument8 pagesWhat'S Inside: Julie PondnwberryfoundationNo ratings yet
- Florence Final Order of DismissalDocument10 pagesFlorence Final Order of DismissalABC15 NewsNo ratings yet
- Ascorbite and CaroteinDocument108 pagesAscorbite and CaroteinfitriasarassNo ratings yet
- Oversight Hearing: Committee On Natural Resources U.S. House of RepresentativesDocument71 pagesOversight Hearing: Committee On Natural Resources U.S. House of RepresentativesScribd Government DocsNo ratings yet
- ERSS Andinoacara Latifrons Final June2018 RevisedDocument9 pagesERSS Andinoacara Latifrons Final June2018 Revisedzaira.890211No ratings yet
- 32 Sfu 8-12-20Document10 pages32 Sfu 8-12-20nwberryfoundationNo ratings yet
- Oversight Hearing: Committee On Natural Resources U.S. House of RepresentativesDocument71 pagesOversight Hearing: Committee On Natural Resources U.S. House of RepresentativesScribd Government DocsNo ratings yet
- Aquaculture-A Gateway For Exotic SpeciesDocument2 pagesAquaculture-A Gateway For Exotic SpeciesgiuseppegnrNo ratings yet
- House Hearing, 108TH Congress - Examining Impacts of The Endangered Species Act On Southern California's Inland EmpireDocument76 pagesHouse Hearing, 108TH Congress - Examining Impacts of The Endangered Species Act On Southern California's Inland EmpireScribd Government DocsNo ratings yet
- Isoflavone Levels, Nodulation and Gene Expression Profiles of A CRISPR/ Cas9 Deletion Mutant in The Isoflavone Synthase Gene of Red CloverDocument1 pageIsoflavone Levels, Nodulation and Gene Expression Profiles of A CRISPR/ Cas9 Deletion Mutant in The Isoflavone Synthase Gene of Red CloversabaNo ratings yet
- 9716 Edwards 2004 Occ 11 1 156Document168 pages9716 Edwards 2004 Occ 11 1 156Moisés Benites DíazNo ratings yet
- JasonDocument7 pagesJasonapi-401676284No ratings yet
- TPG 9 3 Plantgenome2016.01.0009 PDFDocument10 pagesTPG 9 3 Plantgenome2016.01.0009 PDFOanaNo ratings yet
- A Classification of North American Biotic Communities - Brown, Reichenbacher, FransonDocument298 pagesA Classification of North American Biotic Communities - Brown, Reichenbacher, Fransonlihelih1No ratings yet
- Guest Commentary: What'S InsideDocument12 pagesGuest Commentary: What'S InsidenwberryfoundationNo ratings yet
- Carnivorousplantnewsletter 2005Document32 pagesCarnivorousplantnewsletter 2005gppgpp1993No ratings yet
- Week 35: Northwest Berry FoundationDocument5 pagesWeek 35: Northwest Berry FoundationnwberryfoundationNo ratings yet
- NB-80-001 Article-90439 en 1Document119 pagesNB-80-001 Article-90439 en 1santiagoformadorNo ratings yet
- Shewanella and Description of Shewanella Oneidensis Sp. Now: Polyphasic Taxonomy of The GenusDocument20 pagesShewanella and Description of Shewanella Oneidensis Sp. Now: Polyphasic Taxonomy of The Genuserfawn9No ratings yet
- Passport Data - VIVC Prime Name - SUFFOLK REDDocument2 pagesPassport Data - VIVC Prime Name - SUFFOLK REDfafaNo ratings yet
- (ACS Symposium Series 208) Paul A. Hedin (Eds.) - Plant Resistance To Insects-American Chemical Society (1983)Document363 pages(ACS Symposium Series 208) Paul A. Hedin (Eds.) - Plant Resistance To Insects-American Chemical Society (1983)ezhilmaniNo ratings yet
- House Hearing, 114TH Congress - State, Foreign Operations, and Related Programs Appropriations For 2017Document519 pagesHouse Hearing, 114TH Congress - State, Foreign Operations, and Related Programs Appropriations For 2017Scribd Government DocsNo ratings yet
- 34 Sfu 8-26-20Document9 pages34 Sfu 8-26-20nwberryfoundationNo ratings yet
- Amphibian Medicine and Captive Husbandry (VetBooks - Ir)Document555 pagesAmphibian Medicine and Captive Husbandry (VetBooks - Ir)Thalia SantiagoNo ratings yet
- Royal Botanic Gardens, Kew, Springer Kew BulletinDocument11 pagesRoyal Botanic Gardens, Kew, Springer Kew BulletinRosy RodriguesNo ratings yet
- Justice and Natural Resources: Concepts, Strategies, and ApplicationsFrom EverandJustice and Natural Resources: Concepts, Strategies, and ApplicationsRating: 3 out of 5 stars3/5 (1)
- United States District Court For The Northern District of CaliforniaDocument23 pagesUnited States District Court For The Northern District of CaliforniaEquality Case FilesNo ratings yet
- Verticillium Wilt in Tennessee Valley CottonDocument7 pagesVerticillium Wilt in Tennessee Valley CottonCedrus CedrusNo ratings yet
- Alfalfa Seed ProductionDocument17 pagesAlfalfa Seed ProductionGhazali AzmiNo ratings yet
- The Development of Two Flanking SCAR Markers Linked To A Sex Determination Locus in Salix Viminalis LDocument5 pagesThe Development of Two Flanking SCAR Markers Linked To A Sex Determination Locus in Salix Viminalis LpbrasicaNo ratings yet
- Recent Moon Jelly Blooms in KoreanDocument11 pagesRecent Moon Jelly Blooms in KoreanyangNo ratings yet
- Genetic and Morphological Variability in South American RodentDocument13 pagesGenetic and Morphological Variability in South American RodentpomajoluNo ratings yet
- Jurnal KarsinologiDocument6 pagesJurnal KarsinologiRani Eva DewiNo ratings yet
- The Oligochaeta (Annelida) Fauna of Aksu Stream (Antalya)Document8 pagesThe Oligochaeta (Annelida) Fauna of Aksu Stream (Antalya)Rani Eva DewiNo ratings yet
- Bioactivity of Trypsin Inhibitors From Sesame Seeds To Control Plodia Interpunctella Larvae (Hübner) (Lepidoptera: Pyralidae)Document8 pagesBioactivity of Trypsin Inhibitors From Sesame Seeds To Control Plodia Interpunctella Larvae (Hübner) (Lepidoptera: Pyralidae)Rani Eva DewiNo ratings yet
- Jpids Pis043 FullDocument3 pagesJpids Pis043 FullRani Eva DewiNo ratings yet
- 1.farah Mastura Noor AzmanDocument18 pages1.farah Mastura Noor Azmanzulikah89No ratings yet
- Diagnostic Accuracy of The 3-Meter Backward Walk Test in Persons With Parkinson DiseaseDocument6 pagesDiagnostic Accuracy of The 3-Meter Backward Walk Test in Persons With Parkinson DiseaseNITISHA GUPTANo ratings yet
- Regression AnalysisDocument280 pagesRegression AnalysisA.Benhari100% (1)
- Development of A Self-Report Instrument To Measure Patient Safety Attitudes, Skills, and KnowledgeDocument5 pagesDevelopment of A Self-Report Instrument To Measure Patient Safety Attitudes, Skills, and KnowledgeEko Wahyu AgustinNo ratings yet
- Endterm - 1Document14 pagesEndterm - 1himanshubahmaniNo ratings yet
- Designing A Good Instructional Supervisory PlanDocument26 pagesDesigning A Good Instructional Supervisory PlanDENNIS RAMIREZNo ratings yet
- Week 3 Standard Consistency, Setting Time & Fineness of CementDocument6 pagesWeek 3 Standard Consistency, Setting Time & Fineness of CementFareez SedakaNo ratings yet
- Weekly Schedule of MBA 2016 SIIIA Week 1Document12 pagesWeekly Schedule of MBA 2016 SIIIA Week 1aadinNo ratings yet
- Preferred Future of Filipino School Leadership PDFDocument12 pagesPreferred Future of Filipino School Leadership PDFRaiza Basatan CanillasNo ratings yet
- Bneo BGPMSDocument26 pagesBneo BGPMSCarlo TorresNo ratings yet
- Critical Success Factors of Erp Implementation in Pakistan.Document8 pagesCritical Success Factors of Erp Implementation in Pakistan.Tarik MehmoodNo ratings yet
- Hotel Management Thesis SampleDocument6 pagesHotel Management Thesis Samplebk1svxmr100% (2)
- Avelino Et Al - 2019 - Transformative Social InnovationDocument12 pagesAvelino Et Al - 2019 - Transformative Social InnovationJessica PerezNo ratings yet
- Disertasyon Abstract NG SampleDocument11 pagesDisertasyon Abstract NG Samplefloresjen_1450% (2)
- Graham Et Al 2019 The Validity and Reliability of Available Intimate Partner Homicide and Reassault Risk AssessmentDocument23 pagesGraham Et Al 2019 The Validity and Reliability of Available Intimate Partner Homicide and Reassault Risk AssessmentAndrea Guerrero ZapataNo ratings yet
- Chapter-II-METHODS.-ALA-LAZARO (Edited)Document5 pagesChapter-II-METHODS.-ALA-LAZARO (Edited)Princess CandaleraNo ratings yet
- SafetrekbookDocument11 pagesSafetrekbookapi-242471392No ratings yet
- Succession in Family Business: A Review of The ResearchDocument26 pagesSuccession in Family Business: A Review of The ResearchAna Flávia PortezanNo ratings yet
- Impact of Internet Banking On The Customer Satisfaction: Evidence From The Indian Banking SectorDocument9 pagesImpact of Internet Banking On The Customer Satisfaction: Evidence From The Indian Banking SectorKavya MudhirajNo ratings yet
- 1 s2.0 S0386111219301682 MainDocument10 pages1 s2.0 S0386111219301682 Mainquang duNo ratings yet
- Hydro Mechanical Systems in Simulink in SL Part 2 PDFDocument20 pagesHydro Mechanical Systems in Simulink in SL Part 2 PDFAbdulhmeed MutalatNo ratings yet
- Scott Reuschel TANDEM LABS - Regulated Bioanlysis For The Pharmaceutical IndustryDocument33 pagesScott Reuschel TANDEM LABS - Regulated Bioanlysis For The Pharmaceutical IndustrynikuNo ratings yet
- SDLC - Ws-Commonly Asked Interview Questions With AnswersDocument4 pagesSDLC - Ws-Commonly Asked Interview Questions With Answersdhaval88No ratings yet
- Booklet No09 Machine and Process Capability - en PDFDocument108 pagesBooklet No09 Machine and Process Capability - en PDFHari KnowNo ratings yet
- CHAPTER 3 Multiple Linear RegressionDocument35 pagesCHAPTER 3 Multiple Linear RegressionFakhrulShahrilEzanieNo ratings yet
- Characteristics of Brand NamesDocument12 pagesCharacteristics of Brand NamesAhmed AmrNo ratings yet
- Freelance Market Research Analyst 1110Document4 pagesFreelance Market Research Analyst 1110IshanSaneNo ratings yet




























































































Download as pdf or txt
You might also like
- Colorado Flora: Eastern Slope, Fourth Edition A Field Guide to the Vascular PlantsFrom EverandColorado Flora: Eastern Slope, Fourth Edition A Field Guide to the Vascular PlantsRating: 3.5 out of 5 stars3.5/5 (2)
- Power of GravityDocument71 pagesPower of GravityJose RodriguezNo ratings yet
- Adjuvants For Agrichemicals PDFDocument764 pagesAdjuvants For Agrichemicals PDFAlvaro ProañoNo ratings yet
- Ua School of Journalism Professional Project Proposal Guidelines For The Master'S Degree ProgramDocument8 pagesUa School of Journalism Professional Project Proposal Guidelines For The Master'S Degree ProgramChaii Madlangsakay TolentinoNo ratings yet
- Project Report On LakmeDocument53 pagesProject Report On LakmeOmita Johar69% (13)
- South Florida Vegetable Pest and Disease Hotline For January 3, 2018Document24 pagesSouth Florida Vegetable Pest and Disease Hotline For January 3, 2018Gene McAvoyNo ratings yet
- Insecticide Resistance of Diamondback Moth (Lepidoptera: Plutellidae) in North AmericaDocument9 pagesInsecticide Resistance of Diamondback Moth (Lepidoptera: Plutellidae) in North AmericaFlorencia GorordoNo ratings yet
- 314Document4 pages314Carlos MeirellesNo ratings yet
- Castilleja: A Publication of The Wyoming Native Plant SocietyDocument12 pagesCastilleja: A Publication of The Wyoming Native Plant SocietyWyoming Native Plant SocietyNo ratings yet
- Trout Production - United States Department of AgricultureDocument15 pagesTrout Production - United States Department of AgriculturehighmountainbearNo ratings yet
- Arnett Thomas Skelley Frank 2002Document878 pagesArnett Thomas Skelley Frank 2002raitomi04No ratings yet
- YoungEtAl ChenopodDocument17 pagesYoungEtAl ChenopodCHrisNo ratings yet
- Castilleja: A Publication of The Wyoming Native Plant SocietyDocument12 pagesCastilleja: A Publication of The Wyoming Native Plant SocietyWyoming Native Plant SocietyNo ratings yet
- Fishes of North America Endangered, Threatened, or of Special Concern: 1979Document16 pagesFishes of North America Endangered, Threatened, or of Special Concern: 1979guillermo lopezNo ratings yet
- Cut Flowers and FoliagesFrom EverandCut Flowers and FoliagesJames E. FaustNo ratings yet
- Colorado Flora: Western Slope, Fourth Edition A Field Guide to the Vascular PlantsFrom EverandColorado Flora: Western Slope, Fourth Edition A Field Guide to the Vascular PlantsNo ratings yet
- Castilleja: A Publication of The Wyoming Native Plant SocietyDocument10 pagesCastilleja: A Publication of The Wyoming Native Plant SocietyWyoming Native Plant SocietyNo ratings yet
- CDTX PatentDocument364 pagesCDTX PatentCharles GrossNo ratings yet
- American Beetles, v.2 (Arnett Et Al., 2002)Document881 pagesAmerican Beetles, v.2 (Arnett Et Al., 2002)Paco Ramirez100% (1)
- Scholtz Decapod 2009 PDFDocument22 pagesScholtz Decapod 2009 PDFJaqen H'gharNo ratings yet
- Bringraj Eclipta Alba ECPRDocument3 pagesBringraj Eclipta Alba ECPRRajesh KumarNo ratings yet
- Giant Water Bug in AsiaDocument11 pagesGiant Water Bug in AsiaDũng Hoàng TrầnNo ratings yet
- Special Sego Lily Internet Offer For All Members!: NOV/DEC 2002 NOV/DEC 2002Document8 pagesSpecial Sego Lily Internet Offer For All Members!: NOV/DEC 2002 NOV/DEC 2002Friends of Utah Native Plant SocietyNo ratings yet
- Sove 1993, Vol 18, No 2 PDFDocument124 pagesSove 1993, Vol 18, No 2 PDFanggit prihatnoloNo ratings yet
- Jashs Article p554Document4 pagesJashs Article p554raraNo ratings yet
- Pdis-93-6-0584.pdf PatooDocument9 pagesPdis-93-6-0584.pdf PatooJose Miguel Soto HerediaNo ratings yet
- Gao - Et.al-2023-The Generation of The First Chromosome-Level de Novo Genome Assembly and Development and validation-50KSNParray-SalmonDocument14 pagesGao - Et.al-2023-The Generation of The First Chromosome-Level de Novo Genome Assembly and Development and validation-50KSNParray-SalmonRama BangeraNo ratings yet
- Notice of Release DS5-67 (Soybean)Document3 pagesNotice of Release DS5-67 (Soybean)Rodrigo Vazquez ValdezNo ratings yet
- Botany Labels Final FinalDocument11 pagesBotany Labels Final FinalAlexis MoffNo ratings yet
- What'S Inside: Julie PondDocument8 pagesWhat'S Inside: Julie PondnwberryfoundationNo ratings yet
- Florence Final Order of DismissalDocument10 pagesFlorence Final Order of DismissalABC15 NewsNo ratings yet
- Ascorbite and CaroteinDocument108 pagesAscorbite and CaroteinfitriasarassNo ratings yet
- Oversight Hearing: Committee On Natural Resources U.S. House of RepresentativesDocument71 pagesOversight Hearing: Committee On Natural Resources U.S. House of RepresentativesScribd Government DocsNo ratings yet
- ERSS Andinoacara Latifrons Final June2018 RevisedDocument9 pagesERSS Andinoacara Latifrons Final June2018 Revisedzaira.890211No ratings yet
- 32 Sfu 8-12-20Document10 pages32 Sfu 8-12-20nwberryfoundationNo ratings yet
- Oversight Hearing: Committee On Natural Resources U.S. House of RepresentativesDocument71 pagesOversight Hearing: Committee On Natural Resources U.S. House of RepresentativesScribd Government DocsNo ratings yet
- Aquaculture-A Gateway For Exotic SpeciesDocument2 pagesAquaculture-A Gateway For Exotic SpeciesgiuseppegnrNo ratings yet
- House Hearing, 108TH Congress - Examining Impacts of The Endangered Species Act On Southern California's Inland EmpireDocument76 pagesHouse Hearing, 108TH Congress - Examining Impacts of The Endangered Species Act On Southern California's Inland EmpireScribd Government DocsNo ratings yet
- Isoflavone Levels, Nodulation and Gene Expression Profiles of A CRISPR/ Cas9 Deletion Mutant in The Isoflavone Synthase Gene of Red CloverDocument1 pageIsoflavone Levels, Nodulation and Gene Expression Profiles of A CRISPR/ Cas9 Deletion Mutant in The Isoflavone Synthase Gene of Red CloversabaNo ratings yet
- 9716 Edwards 2004 Occ 11 1 156Document168 pages9716 Edwards 2004 Occ 11 1 156Moisés Benites DíazNo ratings yet
- JasonDocument7 pagesJasonapi-401676284No ratings yet
- TPG 9 3 Plantgenome2016.01.0009 PDFDocument10 pagesTPG 9 3 Plantgenome2016.01.0009 PDFOanaNo ratings yet
- A Classification of North American Biotic Communities - Brown, Reichenbacher, FransonDocument298 pagesA Classification of North American Biotic Communities - Brown, Reichenbacher, Fransonlihelih1No ratings yet
- Guest Commentary: What'S InsideDocument12 pagesGuest Commentary: What'S InsidenwberryfoundationNo ratings yet
- Carnivorousplantnewsletter 2005Document32 pagesCarnivorousplantnewsletter 2005gppgpp1993No ratings yet
- Week 35: Northwest Berry FoundationDocument5 pagesWeek 35: Northwest Berry FoundationnwberryfoundationNo ratings yet
- NB-80-001 Article-90439 en 1Document119 pagesNB-80-001 Article-90439 en 1santiagoformadorNo ratings yet
- Shewanella and Description of Shewanella Oneidensis Sp. Now: Polyphasic Taxonomy of The GenusDocument20 pagesShewanella and Description of Shewanella Oneidensis Sp. Now: Polyphasic Taxonomy of The Genuserfawn9No ratings yet
- Passport Data - VIVC Prime Name - SUFFOLK REDDocument2 pagesPassport Data - VIVC Prime Name - SUFFOLK REDfafaNo ratings yet
- (ACS Symposium Series 208) Paul A. Hedin (Eds.) - Plant Resistance To Insects-American Chemical Society (1983)Document363 pages(ACS Symposium Series 208) Paul A. Hedin (Eds.) - Plant Resistance To Insects-American Chemical Society (1983)ezhilmaniNo ratings yet
- House Hearing, 114TH Congress - State, Foreign Operations, and Related Programs Appropriations For 2017Document519 pagesHouse Hearing, 114TH Congress - State, Foreign Operations, and Related Programs Appropriations For 2017Scribd Government DocsNo ratings yet
- 34 Sfu 8-26-20Document9 pages34 Sfu 8-26-20nwberryfoundationNo ratings yet
- Amphibian Medicine and Captive Husbandry (VetBooks - Ir)Document555 pagesAmphibian Medicine and Captive Husbandry (VetBooks - Ir)Thalia SantiagoNo ratings yet
- Royal Botanic Gardens, Kew, Springer Kew BulletinDocument11 pagesRoyal Botanic Gardens, Kew, Springer Kew BulletinRosy RodriguesNo ratings yet
- Justice and Natural Resources: Concepts, Strategies, and ApplicationsFrom EverandJustice and Natural Resources: Concepts, Strategies, and ApplicationsRating: 3 out of 5 stars3/5 (1)
- United States District Court For The Northern District of CaliforniaDocument23 pagesUnited States District Court For The Northern District of CaliforniaEquality Case FilesNo ratings yet
- Verticillium Wilt in Tennessee Valley CottonDocument7 pagesVerticillium Wilt in Tennessee Valley CottonCedrus CedrusNo ratings yet
- Alfalfa Seed ProductionDocument17 pagesAlfalfa Seed ProductionGhazali AzmiNo ratings yet
- The Development of Two Flanking SCAR Markers Linked To A Sex Determination Locus in Salix Viminalis LDocument5 pagesThe Development of Two Flanking SCAR Markers Linked To A Sex Determination Locus in Salix Viminalis LpbrasicaNo ratings yet
- Recent Moon Jelly Blooms in KoreanDocument11 pagesRecent Moon Jelly Blooms in KoreanyangNo ratings yet
- Genetic and Morphological Variability in South American RodentDocument13 pagesGenetic and Morphological Variability in South American RodentpomajoluNo ratings yet
- Jurnal KarsinologiDocument6 pagesJurnal KarsinologiRani Eva DewiNo ratings yet
- The Oligochaeta (Annelida) Fauna of Aksu Stream (Antalya)Document8 pagesThe Oligochaeta (Annelida) Fauna of Aksu Stream (Antalya)Rani Eva DewiNo ratings yet
- Bioactivity of Trypsin Inhibitors From Sesame Seeds To Control Plodia Interpunctella Larvae (Hübner) (Lepidoptera: Pyralidae)Document8 pagesBioactivity of Trypsin Inhibitors From Sesame Seeds To Control Plodia Interpunctella Larvae (Hübner) (Lepidoptera: Pyralidae)Rani Eva DewiNo ratings yet
- Jpids Pis043 FullDocument3 pagesJpids Pis043 FullRani Eva DewiNo ratings yet
- 1.farah Mastura Noor AzmanDocument18 pages1.farah Mastura Noor Azmanzulikah89No ratings yet
- Diagnostic Accuracy of The 3-Meter Backward Walk Test in Persons With Parkinson DiseaseDocument6 pagesDiagnostic Accuracy of The 3-Meter Backward Walk Test in Persons With Parkinson DiseaseNITISHA GUPTANo ratings yet
- Regression AnalysisDocument280 pagesRegression AnalysisA.Benhari100% (1)
- Development of A Self-Report Instrument To Measure Patient Safety Attitudes, Skills, and KnowledgeDocument5 pagesDevelopment of A Self-Report Instrument To Measure Patient Safety Attitudes, Skills, and KnowledgeEko Wahyu AgustinNo ratings yet
- Endterm - 1Document14 pagesEndterm - 1himanshubahmaniNo ratings yet
- Designing A Good Instructional Supervisory PlanDocument26 pagesDesigning A Good Instructional Supervisory PlanDENNIS RAMIREZNo ratings yet
- Week 3 Standard Consistency, Setting Time & Fineness of CementDocument6 pagesWeek 3 Standard Consistency, Setting Time & Fineness of CementFareez SedakaNo ratings yet
- Weekly Schedule of MBA 2016 SIIIA Week 1Document12 pagesWeekly Schedule of MBA 2016 SIIIA Week 1aadinNo ratings yet
- Preferred Future of Filipino School Leadership PDFDocument12 pagesPreferred Future of Filipino School Leadership PDFRaiza Basatan CanillasNo ratings yet
- Bneo BGPMSDocument26 pagesBneo BGPMSCarlo TorresNo ratings yet
- Critical Success Factors of Erp Implementation in Pakistan.Document8 pagesCritical Success Factors of Erp Implementation in Pakistan.Tarik MehmoodNo ratings yet
- Hotel Management Thesis SampleDocument6 pagesHotel Management Thesis Samplebk1svxmr100% (2)
- Avelino Et Al - 2019 - Transformative Social InnovationDocument12 pagesAvelino Et Al - 2019 - Transformative Social InnovationJessica PerezNo ratings yet
- Disertasyon Abstract NG SampleDocument11 pagesDisertasyon Abstract NG Samplefloresjen_1450% (2)
- Graham Et Al 2019 The Validity and Reliability of Available Intimate Partner Homicide and Reassault Risk AssessmentDocument23 pagesGraham Et Al 2019 The Validity and Reliability of Available Intimate Partner Homicide and Reassault Risk AssessmentAndrea Guerrero ZapataNo ratings yet
- Chapter-II-METHODS.-ALA-LAZARO (Edited)Document5 pagesChapter-II-METHODS.-ALA-LAZARO (Edited)Princess CandaleraNo ratings yet
- SafetrekbookDocument11 pagesSafetrekbookapi-242471392No ratings yet
- Succession in Family Business: A Review of The ResearchDocument26 pagesSuccession in Family Business: A Review of The ResearchAna Flávia PortezanNo ratings yet
- Impact of Internet Banking On The Customer Satisfaction: Evidence From The Indian Banking SectorDocument9 pagesImpact of Internet Banking On The Customer Satisfaction: Evidence From The Indian Banking SectorKavya MudhirajNo ratings yet
- 1 s2.0 S0386111219301682 MainDocument10 pages1 s2.0 S0386111219301682 Mainquang duNo ratings yet
- Hydro Mechanical Systems in Simulink in SL Part 2 PDFDocument20 pagesHydro Mechanical Systems in Simulink in SL Part 2 PDFAbdulhmeed MutalatNo ratings yet
- Scott Reuschel TANDEM LABS - Regulated Bioanlysis For The Pharmaceutical IndustryDocument33 pagesScott Reuschel TANDEM LABS - Regulated Bioanlysis For The Pharmaceutical IndustrynikuNo ratings yet
- SDLC - Ws-Commonly Asked Interview Questions With AnswersDocument4 pagesSDLC - Ws-Commonly Asked Interview Questions With Answersdhaval88No ratings yet
- Booklet No09 Machine and Process Capability - en PDFDocument108 pagesBooklet No09 Machine and Process Capability - en PDFHari KnowNo ratings yet
- CHAPTER 3 Multiple Linear RegressionDocument35 pagesCHAPTER 3 Multiple Linear RegressionFakhrulShahrilEzanieNo ratings yet
- Characteristics of Brand NamesDocument12 pagesCharacteristics of Brand NamesAhmed AmrNo ratings yet
- Freelance Market Research Analyst 1110Document4 pagesFreelance Market Research Analyst 1110IshanSaneNo ratings yet