
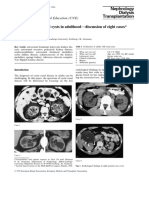
12 Boute 00 NatGenet
12 Boute 00 NatGenet
You might also like
- Pglo Transformation Lab Report - Kierra LeonardDocument9 pagesPglo Transformation Lab Report - Kierra Leonardapi-508539249No ratings yet
- Lab ReportDocument10 pagesLab ReportJeevikaGoyal100% (1)
- NGSDocument252 pagesNGSsaurav choudhary100% (2)
- Genetic Basis of Steroid Resistant Nephrotic Syndrome: Review ArticleDocument7 pagesGenetic Basis of Steroid Resistant Nephrotic Syndrome: Review ArticleIgnacio TabuadaNo ratings yet
- Hereditary Podocytopathies in Adults: The Next Generation: The Kidney in Genetic and Rare Diseases: ReviewDocument7 pagesHereditary Podocytopathies in Adults: The Next Generation: The Kidney in Genetic and Rare Diseases: ReviewIgnacio TabuadaNo ratings yet
- Huber 2001Document4 pagesHuber 2001Flip Flop ChartNo ratings yet
- Polycystic Kidney - Eight CasesDocument11 pagesPolycystic Kidney - Eight CasesKAINAT ANJUMNo ratings yet
- B CatetinDocument20 pagesB Catetinyayu latifahNo ratings yet
- B-Catetin (01-09)Document9 pagesB-Catetin (01-09)yayu latifahNo ratings yet
- Mezzabotta Et Al-2015-Journal of Cellular and Molecular MedicineDocument14 pagesMezzabotta Et Al-2015-Journal of Cellular and Molecular MedicineTania AzhariNo ratings yet
- HHS Public Access: Renal Dysplasia in The NeonateDocument13 pagesHHS Public Access: Renal Dysplasia in The NeonateErwin CandraNo ratings yet
- Nephrotic Syndrome PirDocument15 pagesNephrotic Syndrome PirNosirova ManijaNo ratings yet
- Arch Dis Child 2008 93 (Suppl II) - A47Document1 pageArch Dis Child 2008 93 (Suppl II) - A47Aamir Jalal Al-MosawiNo ratings yet
- 4 UmfDocument5 pages4 UmfAndreea NicoletaNo ratings yet
- EXOSC8Document6 pagesEXOSC8vuhaipham1407No ratings yet
- Icbt 05 I 9 P 777Document9 pagesIcbt 05 I 9 P 777Hj herniatiNo ratings yet
- Assessment of The Distribution of Mitochondrial Ribosomal Rna in Melas and IN Thrombotic Cerebral Infarcts by in HybridizationDocument8 pagesAssessment of The Distribution of Mitochondrial Ribosomal Rna in Melas and IN Thrombotic Cerebral Infarcts by in HybridizationVASCHOTI TRABANICONo ratings yet
- Zehra Agha, Zafar Iqbal, Maleeha Azam, Maimoona Siddique, Marjolein H. Willemsen, Tjitske Kleefstra, Christiane Zweier, Nicole de Leeuw, Raheel Qamar, Hans Van BokhovenDocument6 pagesZehra Agha, Zafar Iqbal, Maleeha Azam, Maimoona Siddique, Marjolein H. Willemsen, Tjitske Kleefstra, Christiane Zweier, Nicole de Leeuw, Raheel Qamar, Hans Van BokhovenAli HaiderNo ratings yet
- Qi-Dan Fang Ameliorates Adriamycin-Induced Nephrotic Syndrome RatDocument9 pagesQi-Dan Fang Ameliorates Adriamycin-Induced Nephrotic Syndrome RatBeatri AyuzaNo ratings yet
- Letters: Whole-Exome Sequencing Identifies Recessive WDR62 Mutations in Severe Brain MalformationsDocument5 pagesLetters: Whole-Exome Sequencing Identifies Recessive WDR62 Mutations in Severe Brain MalformationsAli HaiderNo ratings yet
- Ijms 25 02936 v3Document18 pagesIjms 25 02936 v3Gilson MurataNo ratings yet
- Genetic and Biochemical Background of Chronic Granulomatous DiseaseDocument8 pagesGenetic and Biochemical Background of Chronic Granulomatous Diseaseapi-19976646No ratings yet
- Deciphering Neurodegenerative Diseases Using Long-Read SequencingDocument12 pagesDeciphering Neurodegenerative Diseases Using Long-Read SequencingShavana RajkumarNo ratings yet
- Brain Regional Differences in Hexanucleotide Repeat Lenght in X Linked Dystonia Parkinsonism Using Nanopore SeqDocument9 pagesBrain Regional Differences in Hexanucleotide Repeat Lenght in X Linked Dystonia Parkinsonism Using Nanopore SeqShavana RajkumarNo ratings yet
- Pathologic Classification of Focal Segmental GlomerulosclerosisDocument18 pagesPathologic Classification of Focal Segmental GlomerulosclerosisRhod LadiaNo ratings yet
- Glucosa Afecta A Granule StressDocument21 pagesGlucosa Afecta A Granule Stressb12loaljNo ratings yet
- Neuropharmacology: Sciverse SciencedirectDocument8 pagesNeuropharmacology: Sciverse SciencedirectShawnNo ratings yet
- Noonan Syndrome: Clinical Aspects and Molecular PathogenesisDocument25 pagesNoonan Syndrome: Clinical Aspects and Molecular PathogenesisFlavia ChNo ratings yet
- Endogenous Notch Signaling in Adult Kidneys.13Document17 pagesEndogenous Notch Signaling in Adult Kidneys.13Arista RachmaNo ratings yet
- Biomedicines 12 00080 v2 2Document20 pagesBiomedicines 12 00080 v2 2MarNo ratings yet
- Converging Pathways in Neurodegeneration, From Genetics To MechanismsDocument10 pagesConverging Pathways in Neurodegeneration, From Genetics To MechanismsCony GSNo ratings yet
- Cleavage and Nuclear Translocation of The Caspase 3 Substrate Rho GDP-dissociation Inhibitor, D4-GDI, During ApoptosisDocument8 pagesCleavage and Nuclear Translocation of The Caspase 3 Substrate Rho GDP-dissociation Inhibitor, D4-GDI, During Apoptosisngu trungNo ratings yet
- Ajpcell 00166 2015Document22 pagesAjpcell 00166 2015JasonNo ratings yet
- Chen 2012Document9 pagesChen 2012Aliel MadaniNo ratings yet
- Expression of Advanced Glycation End Products and TheirDocument11 pagesExpression of Advanced Glycation End Products and TheirTamam JauharNo ratings yet
- Diabetes InsipidDocument6 pagesDiabetes InsipidLors CBadiNo ratings yet
- Benzing-2021-Insights Into Glomerular FiltrationDocument10 pagesBenzing-2021-Insights Into Glomerular FiltrationNguyen LequyenNo ratings yet
- 05 Molecular Genetics of The EpilepsiesDocument15 pages05 Molecular Genetics of The EpilepsiesvenkatesannagarajanNo ratings yet
- Nephrotic Syndrome: Practice GapDocument12 pagesNephrotic Syndrome: Practice GapMuhammad Isnaini ZuhriNo ratings yet
- Carroll 2004Document11 pagesCarroll 2004afifahridhahumairahhNo ratings yet
- Nihms317778 PDFDocument10 pagesNihms317778 PDFamraovcinaNo ratings yet
- An Outline of Essential Topics in Glomerular Pathophysiology, Diagnosis, and Treatment For Nephrology TraineesDocument24 pagesAn Outline of Essential Topics in Glomerular Pathophysiology, Diagnosis, and Treatment For Nephrology TraineesKity SarkarNo ratings yet
- DEAD-box RNA helicase Dbp4/DDX10 is an enhancer of α-synuclein toxicity and oligomerizationDocument37 pagesDEAD-box RNA helicase Dbp4/DDX10 is an enhancer of α-synuclein toxicity and oligomerizationdanielNo ratings yet
- Axelrod 2004Document12 pagesAxelrod 2004Nesa An YahdiyaniNo ratings yet
- Oxidative Stress-Induced Circkif18A Downregulation Impairs Mcm7-Mediated Anti-Senescence in Intervertebral Disc DegenerationDocument13 pagesOxidative Stress-Induced Circkif18A Downregulation Impairs Mcm7-Mediated Anti-Senescence in Intervertebral Disc DegenerationValen EstevezNo ratings yet
- The Pathology Associated With Diabetic RetinopathyDocument8 pagesThe Pathology Associated With Diabetic RetinopathyVivi DeviyanaNo ratings yet
- PCKDDocument2 pagesPCKDShams JailaniNo ratings yet
- Insight Into Glomerular Filtration and Albuminuria - NEJMDocument10 pagesInsight Into Glomerular Filtration and Albuminuria - NEJMasalizwa ludlalaNo ratings yet
- JURNALDocument10 pagesJURNALLuses Shantia HaryantoNo ratings yet
- Hepaticka Encefalopatija I ManganDocument12 pagesHepaticka Encefalopatija I ManganIvan MihailovicNo ratings yet
- The Genetic Causes of Nonsyndromic Congenital Retinal Detachment: A Genetic and Phenotypic Study of Pakistani FamiliesDocument9 pagesThe Genetic Causes of Nonsyndromic Congenital Retinal Detachment: A Genetic and Phenotypic Study of Pakistani FamiliesAmilaNo ratings yet
- Biomol SN InvivoDocument11 pagesBiomol SN Invivogalihrahman51No ratings yet
- Adventures in The Pathophysiology of Brain Ischemia: Penumbra, Gene Expression, Neuroprotection The 2002 Thomas Willis Lecture - Myron D. GinsbergDocument11 pagesAdventures in The Pathophysiology of Brain Ischemia: Penumbra, Gene Expression, Neuroprotection The 2002 Thomas Willis Lecture - Myron D. Ginsbergsanti333No ratings yet
- Mda NeuronDocument19 pagesMda NeuronbioladwikoNo ratings yet
- Prion BrainDocument2 pagesPrion BrainD SNo ratings yet
- Aberrant Upregulation of The Glycolytic Enzyme PFKFB3 in CLN7 Neuronal Ceroid LipofuscinosisDocument14 pagesAberrant Upregulation of The Glycolytic Enzyme PFKFB3 in CLN7 Neuronal Ceroid Lipofuscinosisezequiel.grondona.cbaNo ratings yet
- Protein Misfolding and NeurodegenerationDocument6 pagesProtein Misfolding and NeurodegenerationDaniel RomeroNo ratings yet
- Artículo 2Document6 pagesArtículo 2Gabby Hernandez RodriguezNo ratings yet
- JournalDocument18 pagesJournalapi-709221214No ratings yet
- AlbuminuriaDocument10 pagesAlbuminuriaIsrael Armida SodanargNo ratings yet
- Mahajan 2016Document10 pagesMahajan 2016Juan RamirezNo ratings yet
- 10 1016@j Neures 2009 06 004Document12 pages10 1016@j Neures 2009 06 004dr. RiyanNo ratings yet
- The Oxford Handbook of Event-Related Potential Components: U Muc Doi Muci 3 Muci 4 Muci 5 Muci 6 Muci 7 Muci 8 Muci MuciDocument1 pageThe Oxford Handbook of Event-Related Potential Components: U Muc Doi Muci 3 Muci 4 Muci 5 Muci 6 Muci 7 Muci 8 Muci MuciDorin Cristian AntalNo ratings yet
- Clinically Isolated Syndrome (CIS)Document12 pagesClinically Isolated Syndrome (CIS)Dorin Cristian AntalNo ratings yet
- U Muc Doi Muci 3 Muci 4 Muci 5 Muci 6 Muci 7 Muci 8 Muci MuciDocument1 pageU Muc Doi Muci 3 Muci 4 Muci 5 Muci 6 Muci 7 Muci 8 Muci MuciDorin Cristian AntalNo ratings yet
- As A Man Thinketh by James AllenDocument22 pagesAs A Man Thinketh by James AllenDorin Cristian AntalNo ratings yet
- Childhood Nephrotic Syndrome: Steroid Sensitive, Steroid Resistant and BeyondDocument7 pagesChildhood Nephrotic Syndrome: Steroid Sensitive, Steroid Resistant and BeyondDorin Cristian AntalNo ratings yet
- International Registry of Steroid-Resistant Nephrotic Syndrome: Updated Epidemiological and Clinical DataDocument11 pagesInternational Registry of Steroid-Resistant Nephrotic Syndrome: Updated Epidemiological and Clinical DataDorin Cristian AntalNo ratings yet
- Management of Steroid Resistant Nephrotic Syndrome: GuidelinesDocument13 pagesManagement of Steroid Resistant Nephrotic Syndrome: GuidelinesDorin Cristian AntalNo ratings yet
- The Therapeutic Response To Vincristine Was Examined in Seven ChildrenDocument3 pagesThe Therapeutic Response To Vincristine Was Examined in Seven ChildrenDorin Cristian AntalNo ratings yet
- Pattern of Steroid-Resistant Nephrotic Syndrome in Children and The Role of Histopathology: A Single-Centre StudyDocument2 pagesPattern of Steroid-Resistant Nephrotic Syndrome in Children and The Role of Histopathology: A Single-Centre StudyDorin Cristian AntalNo ratings yet
- Management of Steroid Resistant Nephrotic Syndrome: GuidelinesDocument13 pagesManagement of Steroid Resistant Nephrotic Syndrome: GuidelinesDorin Cristian AntalNo ratings yet
- Sesiune An V Seria A Sem Ii S13: Test ORLDocument1 pageSesiune An V Seria A Sem Ii S13: Test ORLDorin Cristian AntalNo ratings yet
- Orar Chirurgie Plastică, Vasculară, Cardiacă - Semestrul IiDocument1 pageOrar Chirurgie Plastică, Vasculară, Cardiacă - Semestrul IiDorin Cristian AntalNo ratings yet
- Icml06TLW ObjectType WolfeDocument4 pagesIcml06TLW ObjectType WolfeDorin Cristian AntalNo ratings yet
- S2HY SlideHandoutsDocument511 pagesS2HY SlideHandoutsDorin Cristian AntalNo ratings yet
- Enzymes Used in RDT Corrected Version EditedDocument43 pagesEnzymes Used in RDT Corrected Version EditedYuppie RajNo ratings yet
- Antibacterial Agents Basis of ActionDocument8 pagesAntibacterial Agents Basis of ActionFrancisco BecerraNo ratings yet
- Bioinformatics DatabaseDocument50 pagesBioinformatics DatabasedhanushumakanthaNo ratings yet
- SNP Basic Background InformationDocument6 pagesSNP Basic Background InformationEamon BarkhordarianNo ratings yet
- AssignmentDocument6 pagesAssignmentsylvia anneNo ratings yet
- Top 50 Global Stem Cell InfluencersDocument25 pagesTop 50 Global Stem Cell InfluencersranasoftNo ratings yet
- Spaces - MU4dMxOmLaVNWfVNvpC PDF ExportDocument58 pagesSpaces - MU4dMxOmLaVNWfVNvpC PDF ExportFaruk YaşNo ratings yet
- Imperial Protein StainDocument1 pageImperial Protein Stainapi-19918078No ratings yet
- Sni 8268-2016Document4 pagesSni 8268-2016Ichsan RamdaniNo ratings yet
- Unit B Enzymes and MetabolismDocument46 pagesUnit B Enzymes and MetabolismShaghulNo ratings yet
- Epi Scheduale EnglishDocument1 pageEpi Scheduale EnglishKasun Senevirathne100% (2)
- Nigeria Plant Quaratine ServiceDocument17 pagesNigeria Plant Quaratine ServicenmadiekeNo ratings yet
- Introduction To Bioinformatics Lab: 10B17BT571 Core Course Credits: 1 L0T0P2Document3 pagesIntroduction To Bioinformatics Lab: 10B17BT571 Core Course Credits: 1 L0T0P2Raj Kumar SoniNo ratings yet
- National Bioeconomy Blueprint USADocument6 pagesNational Bioeconomy Blueprint USAJulieta SeifertNo ratings yet
- Antibodi Monoklonal Dan Protein Sel TunggalDocument18 pagesAntibodi Monoklonal Dan Protein Sel TunggalMeri RahmadaniNo ratings yet
- 71830-Article Text-155973-1-10-20111109 PDFDocument8 pages71830-Article Text-155973-1-10-20111109 PDFMuktanand DhurveNo ratings yet
- Bio 1Document20 pagesBio 1vvm .s3023No ratings yet
- Mrs B MounikaDocument1 pageMrs B MounikaSanjeev KurapatiNo ratings yet
- Chapter 2: (Genetics) : Heredity, Inheritance and VariationDocument11 pagesChapter 2: (Genetics) : Heredity, Inheritance and VariationRonan BartolazoNo ratings yet
- ECB3e Chapter 20Document80 pagesECB3e Chapter 20Sarifa Ghina RaquelleNo ratings yet
- Meselson and Stahl Experiment DeterminedDocument1 pageMeselson and Stahl Experiment DeterminedCarolina RelvaNo ratings yet
- Paper Reading 2Document43 pagesPaper Reading 2anyou.5487.md07No ratings yet
- Structural Changes of ChromosomesDocument14 pagesStructural Changes of Chromosomesf.s.mugdho1No ratings yet
- Medicomama's Biochemistry QBDocument28 pagesMedicomama's Biochemistry QBbhageerathavmc98No ratings yet
- Mark Scheme (Results) January 2018Document25 pagesMark Scheme (Results) January 2018MahmoudAboAlSamenNo ratings yet
- Beacon Designer ManualDocument183 pagesBeacon Designer Manualdnajen100% (1)
- BT503 Environment Biotechnology (Merget PPT Handouts (1 To 235) Full Mid and Final)Document139 pagesBT503 Environment Biotechnology (Merget PPT Handouts (1 To 235) Full Mid and Final)laibaNo ratings yet





































































































Download as pdf or txt
You might also like
- Pglo Transformation Lab Report - Kierra LeonardDocument9 pagesPglo Transformation Lab Report - Kierra Leonardapi-508539249No ratings yet
- Lab ReportDocument10 pagesLab ReportJeevikaGoyal100% (1)
- NGSDocument252 pagesNGSsaurav choudhary100% (2)
- Genetic Basis of Steroid Resistant Nephrotic Syndrome: Review ArticleDocument7 pagesGenetic Basis of Steroid Resistant Nephrotic Syndrome: Review ArticleIgnacio TabuadaNo ratings yet
- Hereditary Podocytopathies in Adults: The Next Generation: The Kidney in Genetic and Rare Diseases: ReviewDocument7 pagesHereditary Podocytopathies in Adults: The Next Generation: The Kidney in Genetic and Rare Diseases: ReviewIgnacio TabuadaNo ratings yet
- Huber 2001Document4 pagesHuber 2001Flip Flop ChartNo ratings yet
- Polycystic Kidney - Eight CasesDocument11 pagesPolycystic Kidney - Eight CasesKAINAT ANJUMNo ratings yet
- B CatetinDocument20 pagesB Catetinyayu latifahNo ratings yet
- B-Catetin (01-09)Document9 pagesB-Catetin (01-09)yayu latifahNo ratings yet
- Mezzabotta Et Al-2015-Journal of Cellular and Molecular MedicineDocument14 pagesMezzabotta Et Al-2015-Journal of Cellular and Molecular MedicineTania AzhariNo ratings yet
- HHS Public Access: Renal Dysplasia in The NeonateDocument13 pagesHHS Public Access: Renal Dysplasia in The NeonateErwin CandraNo ratings yet
- Nephrotic Syndrome PirDocument15 pagesNephrotic Syndrome PirNosirova ManijaNo ratings yet
- Arch Dis Child 2008 93 (Suppl II) - A47Document1 pageArch Dis Child 2008 93 (Suppl II) - A47Aamir Jalal Al-MosawiNo ratings yet
- 4 UmfDocument5 pages4 UmfAndreea NicoletaNo ratings yet
- EXOSC8Document6 pagesEXOSC8vuhaipham1407No ratings yet
- Icbt 05 I 9 P 777Document9 pagesIcbt 05 I 9 P 777Hj herniatiNo ratings yet
- Assessment of The Distribution of Mitochondrial Ribosomal Rna in Melas and IN Thrombotic Cerebral Infarcts by in HybridizationDocument8 pagesAssessment of The Distribution of Mitochondrial Ribosomal Rna in Melas and IN Thrombotic Cerebral Infarcts by in HybridizationVASCHOTI TRABANICONo ratings yet
- Zehra Agha, Zafar Iqbal, Maleeha Azam, Maimoona Siddique, Marjolein H. Willemsen, Tjitske Kleefstra, Christiane Zweier, Nicole de Leeuw, Raheel Qamar, Hans Van BokhovenDocument6 pagesZehra Agha, Zafar Iqbal, Maleeha Azam, Maimoona Siddique, Marjolein H. Willemsen, Tjitske Kleefstra, Christiane Zweier, Nicole de Leeuw, Raheel Qamar, Hans Van BokhovenAli HaiderNo ratings yet
- Qi-Dan Fang Ameliorates Adriamycin-Induced Nephrotic Syndrome RatDocument9 pagesQi-Dan Fang Ameliorates Adriamycin-Induced Nephrotic Syndrome RatBeatri AyuzaNo ratings yet
- Letters: Whole-Exome Sequencing Identifies Recessive WDR62 Mutations in Severe Brain MalformationsDocument5 pagesLetters: Whole-Exome Sequencing Identifies Recessive WDR62 Mutations in Severe Brain MalformationsAli HaiderNo ratings yet
- Ijms 25 02936 v3Document18 pagesIjms 25 02936 v3Gilson MurataNo ratings yet
- Genetic and Biochemical Background of Chronic Granulomatous DiseaseDocument8 pagesGenetic and Biochemical Background of Chronic Granulomatous Diseaseapi-19976646No ratings yet
- Deciphering Neurodegenerative Diseases Using Long-Read SequencingDocument12 pagesDeciphering Neurodegenerative Diseases Using Long-Read SequencingShavana RajkumarNo ratings yet
- Brain Regional Differences in Hexanucleotide Repeat Lenght in X Linked Dystonia Parkinsonism Using Nanopore SeqDocument9 pagesBrain Regional Differences in Hexanucleotide Repeat Lenght in X Linked Dystonia Parkinsonism Using Nanopore SeqShavana RajkumarNo ratings yet
- Pathologic Classification of Focal Segmental GlomerulosclerosisDocument18 pagesPathologic Classification of Focal Segmental GlomerulosclerosisRhod LadiaNo ratings yet
- Glucosa Afecta A Granule StressDocument21 pagesGlucosa Afecta A Granule Stressb12loaljNo ratings yet
- Neuropharmacology: Sciverse SciencedirectDocument8 pagesNeuropharmacology: Sciverse SciencedirectShawnNo ratings yet
- Noonan Syndrome: Clinical Aspects and Molecular PathogenesisDocument25 pagesNoonan Syndrome: Clinical Aspects and Molecular PathogenesisFlavia ChNo ratings yet
- Endogenous Notch Signaling in Adult Kidneys.13Document17 pagesEndogenous Notch Signaling in Adult Kidneys.13Arista RachmaNo ratings yet
- Biomedicines 12 00080 v2 2Document20 pagesBiomedicines 12 00080 v2 2MarNo ratings yet
- Converging Pathways in Neurodegeneration, From Genetics To MechanismsDocument10 pagesConverging Pathways in Neurodegeneration, From Genetics To MechanismsCony GSNo ratings yet
- Cleavage and Nuclear Translocation of The Caspase 3 Substrate Rho GDP-dissociation Inhibitor, D4-GDI, During ApoptosisDocument8 pagesCleavage and Nuclear Translocation of The Caspase 3 Substrate Rho GDP-dissociation Inhibitor, D4-GDI, During Apoptosisngu trungNo ratings yet
- Ajpcell 00166 2015Document22 pagesAjpcell 00166 2015JasonNo ratings yet
- Chen 2012Document9 pagesChen 2012Aliel MadaniNo ratings yet
- Expression of Advanced Glycation End Products and TheirDocument11 pagesExpression of Advanced Glycation End Products and TheirTamam JauharNo ratings yet
- Diabetes InsipidDocument6 pagesDiabetes InsipidLors CBadiNo ratings yet
- Benzing-2021-Insights Into Glomerular FiltrationDocument10 pagesBenzing-2021-Insights Into Glomerular FiltrationNguyen LequyenNo ratings yet
- 05 Molecular Genetics of The EpilepsiesDocument15 pages05 Molecular Genetics of The EpilepsiesvenkatesannagarajanNo ratings yet
- Nephrotic Syndrome: Practice GapDocument12 pagesNephrotic Syndrome: Practice GapMuhammad Isnaini ZuhriNo ratings yet
- Carroll 2004Document11 pagesCarroll 2004afifahridhahumairahhNo ratings yet
- Nihms317778 PDFDocument10 pagesNihms317778 PDFamraovcinaNo ratings yet
- An Outline of Essential Topics in Glomerular Pathophysiology, Diagnosis, and Treatment For Nephrology TraineesDocument24 pagesAn Outline of Essential Topics in Glomerular Pathophysiology, Diagnosis, and Treatment For Nephrology TraineesKity SarkarNo ratings yet
- DEAD-box RNA helicase Dbp4/DDX10 is an enhancer of α-synuclein toxicity and oligomerizationDocument37 pagesDEAD-box RNA helicase Dbp4/DDX10 is an enhancer of α-synuclein toxicity and oligomerizationdanielNo ratings yet
- Axelrod 2004Document12 pagesAxelrod 2004Nesa An YahdiyaniNo ratings yet
- Oxidative Stress-Induced Circkif18A Downregulation Impairs Mcm7-Mediated Anti-Senescence in Intervertebral Disc DegenerationDocument13 pagesOxidative Stress-Induced Circkif18A Downregulation Impairs Mcm7-Mediated Anti-Senescence in Intervertebral Disc DegenerationValen EstevezNo ratings yet
- The Pathology Associated With Diabetic RetinopathyDocument8 pagesThe Pathology Associated With Diabetic RetinopathyVivi DeviyanaNo ratings yet
- PCKDDocument2 pagesPCKDShams JailaniNo ratings yet
- Insight Into Glomerular Filtration and Albuminuria - NEJMDocument10 pagesInsight Into Glomerular Filtration and Albuminuria - NEJMasalizwa ludlalaNo ratings yet
- JURNALDocument10 pagesJURNALLuses Shantia HaryantoNo ratings yet
- Hepaticka Encefalopatija I ManganDocument12 pagesHepaticka Encefalopatija I ManganIvan MihailovicNo ratings yet
- The Genetic Causes of Nonsyndromic Congenital Retinal Detachment: A Genetic and Phenotypic Study of Pakistani FamiliesDocument9 pagesThe Genetic Causes of Nonsyndromic Congenital Retinal Detachment: A Genetic and Phenotypic Study of Pakistani FamiliesAmilaNo ratings yet
- Biomol SN InvivoDocument11 pagesBiomol SN Invivogalihrahman51No ratings yet
- Adventures in The Pathophysiology of Brain Ischemia: Penumbra, Gene Expression, Neuroprotection The 2002 Thomas Willis Lecture - Myron D. GinsbergDocument11 pagesAdventures in The Pathophysiology of Brain Ischemia: Penumbra, Gene Expression, Neuroprotection The 2002 Thomas Willis Lecture - Myron D. Ginsbergsanti333No ratings yet
- Mda NeuronDocument19 pagesMda NeuronbioladwikoNo ratings yet
- Prion BrainDocument2 pagesPrion BrainD SNo ratings yet
- Aberrant Upregulation of The Glycolytic Enzyme PFKFB3 in CLN7 Neuronal Ceroid LipofuscinosisDocument14 pagesAberrant Upregulation of The Glycolytic Enzyme PFKFB3 in CLN7 Neuronal Ceroid Lipofuscinosisezequiel.grondona.cbaNo ratings yet
- Protein Misfolding and NeurodegenerationDocument6 pagesProtein Misfolding and NeurodegenerationDaniel RomeroNo ratings yet
- Artículo 2Document6 pagesArtículo 2Gabby Hernandez RodriguezNo ratings yet
- JournalDocument18 pagesJournalapi-709221214No ratings yet
- AlbuminuriaDocument10 pagesAlbuminuriaIsrael Armida SodanargNo ratings yet
- Mahajan 2016Document10 pagesMahajan 2016Juan RamirezNo ratings yet
- 10 1016@j Neures 2009 06 004Document12 pages10 1016@j Neures 2009 06 004dr. RiyanNo ratings yet
- The Oxford Handbook of Event-Related Potential Components: U Muc Doi Muci 3 Muci 4 Muci 5 Muci 6 Muci 7 Muci 8 Muci MuciDocument1 pageThe Oxford Handbook of Event-Related Potential Components: U Muc Doi Muci 3 Muci 4 Muci 5 Muci 6 Muci 7 Muci 8 Muci MuciDorin Cristian AntalNo ratings yet
- Clinically Isolated Syndrome (CIS)Document12 pagesClinically Isolated Syndrome (CIS)Dorin Cristian AntalNo ratings yet
- U Muc Doi Muci 3 Muci 4 Muci 5 Muci 6 Muci 7 Muci 8 Muci MuciDocument1 pageU Muc Doi Muci 3 Muci 4 Muci 5 Muci 6 Muci 7 Muci 8 Muci MuciDorin Cristian AntalNo ratings yet
- As A Man Thinketh by James AllenDocument22 pagesAs A Man Thinketh by James AllenDorin Cristian AntalNo ratings yet
- Childhood Nephrotic Syndrome: Steroid Sensitive, Steroid Resistant and BeyondDocument7 pagesChildhood Nephrotic Syndrome: Steroid Sensitive, Steroid Resistant and BeyondDorin Cristian AntalNo ratings yet
- International Registry of Steroid-Resistant Nephrotic Syndrome: Updated Epidemiological and Clinical DataDocument11 pagesInternational Registry of Steroid-Resistant Nephrotic Syndrome: Updated Epidemiological and Clinical DataDorin Cristian AntalNo ratings yet
- Management of Steroid Resistant Nephrotic Syndrome: GuidelinesDocument13 pagesManagement of Steroid Resistant Nephrotic Syndrome: GuidelinesDorin Cristian AntalNo ratings yet
- The Therapeutic Response To Vincristine Was Examined in Seven ChildrenDocument3 pagesThe Therapeutic Response To Vincristine Was Examined in Seven ChildrenDorin Cristian AntalNo ratings yet
- Pattern of Steroid-Resistant Nephrotic Syndrome in Children and The Role of Histopathology: A Single-Centre StudyDocument2 pagesPattern of Steroid-Resistant Nephrotic Syndrome in Children and The Role of Histopathology: A Single-Centre StudyDorin Cristian AntalNo ratings yet
- Management of Steroid Resistant Nephrotic Syndrome: GuidelinesDocument13 pagesManagement of Steroid Resistant Nephrotic Syndrome: GuidelinesDorin Cristian AntalNo ratings yet
- Sesiune An V Seria A Sem Ii S13: Test ORLDocument1 pageSesiune An V Seria A Sem Ii S13: Test ORLDorin Cristian AntalNo ratings yet
- Orar Chirurgie Plastică, Vasculară, Cardiacă - Semestrul IiDocument1 pageOrar Chirurgie Plastică, Vasculară, Cardiacă - Semestrul IiDorin Cristian AntalNo ratings yet
- Icml06TLW ObjectType WolfeDocument4 pagesIcml06TLW ObjectType WolfeDorin Cristian AntalNo ratings yet
- S2HY SlideHandoutsDocument511 pagesS2HY SlideHandoutsDorin Cristian AntalNo ratings yet
- Enzymes Used in RDT Corrected Version EditedDocument43 pagesEnzymes Used in RDT Corrected Version EditedYuppie RajNo ratings yet
- Antibacterial Agents Basis of ActionDocument8 pagesAntibacterial Agents Basis of ActionFrancisco BecerraNo ratings yet
- Bioinformatics DatabaseDocument50 pagesBioinformatics DatabasedhanushumakanthaNo ratings yet
- SNP Basic Background InformationDocument6 pagesSNP Basic Background InformationEamon BarkhordarianNo ratings yet
- AssignmentDocument6 pagesAssignmentsylvia anneNo ratings yet
- Top 50 Global Stem Cell InfluencersDocument25 pagesTop 50 Global Stem Cell InfluencersranasoftNo ratings yet
- Spaces - MU4dMxOmLaVNWfVNvpC PDF ExportDocument58 pagesSpaces - MU4dMxOmLaVNWfVNvpC PDF ExportFaruk YaşNo ratings yet
- Imperial Protein StainDocument1 pageImperial Protein Stainapi-19918078No ratings yet
- Sni 8268-2016Document4 pagesSni 8268-2016Ichsan RamdaniNo ratings yet
- Unit B Enzymes and MetabolismDocument46 pagesUnit B Enzymes and MetabolismShaghulNo ratings yet
- Epi Scheduale EnglishDocument1 pageEpi Scheduale EnglishKasun Senevirathne100% (2)
- Nigeria Plant Quaratine ServiceDocument17 pagesNigeria Plant Quaratine ServicenmadiekeNo ratings yet
- Introduction To Bioinformatics Lab: 10B17BT571 Core Course Credits: 1 L0T0P2Document3 pagesIntroduction To Bioinformatics Lab: 10B17BT571 Core Course Credits: 1 L0T0P2Raj Kumar SoniNo ratings yet
- National Bioeconomy Blueprint USADocument6 pagesNational Bioeconomy Blueprint USAJulieta SeifertNo ratings yet
- Antibodi Monoklonal Dan Protein Sel TunggalDocument18 pagesAntibodi Monoklonal Dan Protein Sel TunggalMeri RahmadaniNo ratings yet
- 71830-Article Text-155973-1-10-20111109 PDFDocument8 pages71830-Article Text-155973-1-10-20111109 PDFMuktanand DhurveNo ratings yet
- Bio 1Document20 pagesBio 1vvm .s3023No ratings yet
- Mrs B MounikaDocument1 pageMrs B MounikaSanjeev KurapatiNo ratings yet
- Chapter 2: (Genetics) : Heredity, Inheritance and VariationDocument11 pagesChapter 2: (Genetics) : Heredity, Inheritance and VariationRonan BartolazoNo ratings yet
- ECB3e Chapter 20Document80 pagesECB3e Chapter 20Sarifa Ghina RaquelleNo ratings yet
- Meselson and Stahl Experiment DeterminedDocument1 pageMeselson and Stahl Experiment DeterminedCarolina RelvaNo ratings yet
- Paper Reading 2Document43 pagesPaper Reading 2anyou.5487.md07No ratings yet
- Structural Changes of ChromosomesDocument14 pagesStructural Changes of Chromosomesf.s.mugdho1No ratings yet
- Medicomama's Biochemistry QBDocument28 pagesMedicomama's Biochemistry QBbhageerathavmc98No ratings yet
- Mark Scheme (Results) January 2018Document25 pagesMark Scheme (Results) January 2018MahmoudAboAlSamenNo ratings yet
- Beacon Designer ManualDocument183 pagesBeacon Designer Manualdnajen100% (1)
- BT503 Environment Biotechnology (Merget PPT Handouts (1 To 235) Full Mid and Final)Document139 pagesBT503 Environment Biotechnology (Merget PPT Handouts (1 To 235) Full Mid and Final)laibaNo ratings yet