
Fysiologie Van Het Zenuwstelsel
Fysiologie Van Het Zenuwstelsel
You might also like
- Samenvatting Blok 1.1 - Groei & OntwikkelingDocument97 pagesSamenvatting Blok 1.1 - Groei & OntwikkelingJasper LaumenNo ratings yet
- VsehDocument158 pagesVsehFloor VerbeekNo ratings yet
- Open Biologie H18, H19 & H20 Samenvatting VWO4Document29 pagesOpen Biologie H18, H19 & H20 Samenvatting VWO4Mynt FNo ratings yet
- Vanders Hoofdstuk 7Document3 pagesVanders Hoofdstuk 7Adolfo SanchezNo ratings yet
- Natuurwetenschappen GO 2 Thema 1 InkijkexemplDocument31 pagesNatuurwetenschappen GO 2 Thema 1 InkijkexemplValerie VillacortaNo ratings yet
- Samenvatting Dwarslaesie RevalidatieDocument9 pagesSamenvatting Dwarslaesie RevalidatieSanne EngelNo ratings yet
- Actieve en Passieve StabiliteitDocument6 pagesActieve en Passieve StabiliteitSabine Van T VeldNo ratings yet
- Welke Soorten Spierweefsels Zijn Er Casus 7Document4 pagesWelke Soorten Spierweefsels Zijn Er Casus 7douniataouachNo ratings yet
- Samenvatting OBDocument27 pagesSamenvatting OBMaaike van BovenNo ratings yet
- Anatomie, Fysiologie en Onderzoek Van Het Vestibulaire SysteemDocument54 pagesAnatomie, Fysiologie en Onderzoek Van Het Vestibulaire SysteemAndreea L. MihalceaNo ratings yet
- Biologie Zusammenfassung h19 + h21Document5 pagesBiologie Zusammenfassung h19 + h21Ilayda AydoganNo ratings yet
- Biopagina UitlegDocument11 pagesBiopagina UitlegtamaraNo ratings yet
- Neuro WoordenlijstDocument9 pagesNeuro Woordenlijstisaline.guedonNo ratings yet
- Biologie Hst. 1 Regeling SamenvattingDocument9 pagesBiologie Hst. 1 Regeling Samenvattingmarealtena06No ratings yet
- Spierfunctie MafDocument5 pagesSpierfunctie MafnenitjeNo ratings yet
- HC Anatomie en FysiologieDocument80 pagesHC Anatomie en FysiologiephillykolenNo ratings yet
- Fysiologie Maag-DarmKanaalDocument31 pagesFysiologie Maag-DarmKanaalFloor VerbeekNo ratings yet
- Het SpierstelselDocument14 pagesHet SpierstelselHilde CampNo ratings yet
- Samenvatting Biologie Voor Jouw Hoofdstuk 1 5VWODocument10 pagesSamenvatting Biologie Voor Jouw Hoofdstuk 1 5VWOblueframedwindowNo ratings yet
- Week 3 College AantekeningenDocument5 pagesWeek 3 College AantekeningenLuna SartoriNo ratings yet
- Samenvatting Vorm en Functie Relaties PDFDocument28 pagesSamenvatting Vorm en Functie Relaties PDFRianne RijkenNo ratings yet
- Samenvatting Kennistoets 3.1: ZoederuiterDocument61 pagesSamenvatting Kennistoets 3.1: ZoederuiterAnne OuweneelNo ratings yet
- Loco Tutorial Vraag 1Document3 pagesLoco Tutorial Vraag 1Tybault Hollanders100% (1)
- Samenvatting Interactie Op CelniveauDocument11 pagesSamenvatting Interactie Op CelniveauPantea PanahiNo ratings yet
- Tentamen Periode 1Document24 pagesTentamen Periode 1benjischilder98No ratings yet
- Cardiovasculair Systeem DutchDocument39 pagesCardiovasculair Systeem DutchHovanes ArakelianNo ratings yet
- Antwoorden HGDocument11 pagesAntwoorden HGnikitadalennceNo ratings yet
- Kopie Van Samenvatting 14.1 BiologieDocument10 pagesKopie Van Samenvatting 14.1 BiologieAnnemijn AnnemijnNo ratings yet
- Samenvatting Biologische PsychologieDocument39 pagesSamenvatting Biologische PsychologieNathalie MoonsNo ratings yet
- Practica Samenvatting OudDocument36 pagesPractica Samenvatting OudOscar van HengelNo ratings yet
- 2 1 4Document14 pages2 1 4Selami YildirmazNo ratings yet
- Samenvatting Hijdra DineshGDocument70 pagesSamenvatting Hijdra DineshGDinesh GokoelNo ratings yet
- Biologie Thema 5 Boek 5b Deel 2Document8 pagesBiologie Thema 5 Boek 5b Deel 2Dima NajiNo ratings yet
- Examen Biologie JuniDocument18 pagesExamen Biologie JuniIsmaïl BerdaïNo ratings yet
- Het ZENUWSTELSEL 3Document39 pagesHet ZENUWSTELSEL 3angeliqueboekelman100% (2)
- Hoofdstuk 7Document14 pagesHoofdstuk 7Navepreet KaurNo ratings yet
- SpiereDocument6 pagesSpiereLilithNo ratings yet
- Humane Embryogenesis 2Document9 pagesHumane Embryogenesis 2ViktoriaNo ratings yet
- TUTM Week 2Document9 pagesTUTM Week 2Jelte FolkertsmaNo ratings yet
- FulltextDocument111 pagesFulltextJoepKluwenNo ratings yet
- 1B1 Week 1 HC4 Ionkanalen en HartpotentialenDocument10 pages1B1 Week 1 HC4 Ionkanalen en HartpotentialenVera VoogtNo ratings yet
- UntitledDocument2 pagesUntitledMichiel KoolenNo ratings yet
- Anatomie Hersenen PDFDocument3 pagesAnatomie Hersenen PDFJulia WindhorstNo ratings yet
- Brein Samenvatting PDFDocument50 pagesBrein Samenvatting PDFAmy SavageNo ratings yet
- Piramide BanenDocument5 pagesPiramide BanendiederikNo ratings yet
- Neuroanatomie 1Document127 pagesNeuroanatomie 1Tamar FoppenNo ratings yet
- Ar Thro LogieDocument23 pagesAr Thro LogielieselvandermaesNo ratings yet
- Deel 3 Hoofdstukken 1-3Document20 pagesDeel 3 Hoofdstukken 1-3Bram GeubbelmansNo ratings yet
- An atDocument7 pagesAn atGosse KrikkeNo ratings yet
- Leerdoelen Taak 2Document8 pagesLeerdoelen Taak 2Bram WerneNo ratings yet
- Anatomie NederlandDocument67 pagesAnatomie NederlandTom StevensNo ratings yet
- Tut MDocument8 pagesTut MRaoul van der PloegNo ratings yet
- Biologie Samenvatting Hoofdstuk 1Document4 pagesBiologie Samenvatting Hoofdstuk 1Isa GroenenNo ratings yet
- Biologie SamenvattingDocument5 pagesBiologie SamenvattingValeria MolNo ratings yet
- Orgaansystemen PDFDocument48 pagesOrgaansystemen PDFMichiel HentenaarNo ratings yet
- Antwoorden InstaptoetsenDocument3 pagesAntwoorden InstaptoetsenLevi WNo ratings yet
- AtrokinematiecaDocument3 pagesAtrokinematiecaSabine Van T VeldNo ratings yet
- Belangrijk MSA KTDocument15 pagesBelangrijk MSA KTTessa HodesNo ratings yet
- SV Waarneming VWO2 Biologie Voor JouDocument8 pagesSV Waarneming VWO2 Biologie Voor JoujuliaNo ratings yet
- Embryologie Samenvatting HC 1-5Document14 pagesEmbryologie Samenvatting HC 1-5Robert-Jan ArkeNo ratings yet
- Samenvatting StatistiekDocument6 pagesSamenvatting StatistiekFloor VerbeekNo ratings yet
- Samenvatting Interne Geneeskunde Hoofdstuk 22Document25 pagesSamenvatting Interne Geneeskunde Hoofdstuk 22Floor VerbeekNo ratings yet
- Spierstelsel Onderbeen en VoetDocument31 pagesSpierstelsel Onderbeen en VoetFloor VerbeekNo ratings yet
- Fysiologie Maag-DarmKanaalDocument31 pagesFysiologie Maag-DarmKanaalFloor VerbeekNo ratings yet
- Histologie en Celbiologie MDLDocument19 pagesHistologie en Celbiologie MDLFloor VerbeekNo ratings yet
- Basisbegrippen FarmacologieDocument10 pagesBasisbegrippen FarmacologieFloor VerbeekNo ratings yet
- Anatomie Van Het BovenbeenDocument13 pagesAnatomie Van Het BovenbeenFloor VerbeekNo ratings yet
- Anatomie Van de HeupDocument13 pagesAnatomie Van de HeupFloor VerbeekNo ratings yet








































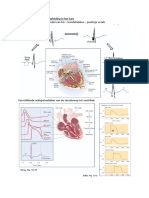







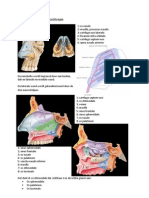


















Download as docx, pdf, or txt
You might also like
- Samenvatting Blok 1.1 - Groei & OntwikkelingDocument97 pagesSamenvatting Blok 1.1 - Groei & OntwikkelingJasper LaumenNo ratings yet
- VsehDocument158 pagesVsehFloor VerbeekNo ratings yet
- Open Biologie H18, H19 & H20 Samenvatting VWO4Document29 pagesOpen Biologie H18, H19 & H20 Samenvatting VWO4Mynt FNo ratings yet
- Vanders Hoofdstuk 7Document3 pagesVanders Hoofdstuk 7Adolfo SanchezNo ratings yet
- Natuurwetenschappen GO 2 Thema 1 InkijkexemplDocument31 pagesNatuurwetenschappen GO 2 Thema 1 InkijkexemplValerie VillacortaNo ratings yet
- Samenvatting Dwarslaesie RevalidatieDocument9 pagesSamenvatting Dwarslaesie RevalidatieSanne EngelNo ratings yet
- Actieve en Passieve StabiliteitDocument6 pagesActieve en Passieve StabiliteitSabine Van T VeldNo ratings yet
- Welke Soorten Spierweefsels Zijn Er Casus 7Document4 pagesWelke Soorten Spierweefsels Zijn Er Casus 7douniataouachNo ratings yet
- Samenvatting OBDocument27 pagesSamenvatting OBMaaike van BovenNo ratings yet
- Anatomie, Fysiologie en Onderzoek Van Het Vestibulaire SysteemDocument54 pagesAnatomie, Fysiologie en Onderzoek Van Het Vestibulaire SysteemAndreea L. MihalceaNo ratings yet
- Biologie Zusammenfassung h19 + h21Document5 pagesBiologie Zusammenfassung h19 + h21Ilayda AydoganNo ratings yet
- Biopagina UitlegDocument11 pagesBiopagina UitlegtamaraNo ratings yet
- Neuro WoordenlijstDocument9 pagesNeuro Woordenlijstisaline.guedonNo ratings yet
- Biologie Hst. 1 Regeling SamenvattingDocument9 pagesBiologie Hst. 1 Regeling Samenvattingmarealtena06No ratings yet
- Spierfunctie MafDocument5 pagesSpierfunctie MafnenitjeNo ratings yet
- HC Anatomie en FysiologieDocument80 pagesHC Anatomie en FysiologiephillykolenNo ratings yet
- Fysiologie Maag-DarmKanaalDocument31 pagesFysiologie Maag-DarmKanaalFloor VerbeekNo ratings yet
- Het SpierstelselDocument14 pagesHet SpierstelselHilde CampNo ratings yet
- Samenvatting Biologie Voor Jouw Hoofdstuk 1 5VWODocument10 pagesSamenvatting Biologie Voor Jouw Hoofdstuk 1 5VWOblueframedwindowNo ratings yet
- Week 3 College AantekeningenDocument5 pagesWeek 3 College AantekeningenLuna SartoriNo ratings yet
- Samenvatting Vorm en Functie Relaties PDFDocument28 pagesSamenvatting Vorm en Functie Relaties PDFRianne RijkenNo ratings yet
- Samenvatting Kennistoets 3.1: ZoederuiterDocument61 pagesSamenvatting Kennistoets 3.1: ZoederuiterAnne OuweneelNo ratings yet
- Loco Tutorial Vraag 1Document3 pagesLoco Tutorial Vraag 1Tybault Hollanders100% (1)
- Samenvatting Interactie Op CelniveauDocument11 pagesSamenvatting Interactie Op CelniveauPantea PanahiNo ratings yet
- Tentamen Periode 1Document24 pagesTentamen Periode 1benjischilder98No ratings yet
- Cardiovasculair Systeem DutchDocument39 pagesCardiovasculair Systeem DutchHovanes ArakelianNo ratings yet
- Antwoorden HGDocument11 pagesAntwoorden HGnikitadalennceNo ratings yet
- Kopie Van Samenvatting 14.1 BiologieDocument10 pagesKopie Van Samenvatting 14.1 BiologieAnnemijn AnnemijnNo ratings yet
- Samenvatting Biologische PsychologieDocument39 pagesSamenvatting Biologische PsychologieNathalie MoonsNo ratings yet
- Practica Samenvatting OudDocument36 pagesPractica Samenvatting OudOscar van HengelNo ratings yet
- 2 1 4Document14 pages2 1 4Selami YildirmazNo ratings yet
- Samenvatting Hijdra DineshGDocument70 pagesSamenvatting Hijdra DineshGDinesh GokoelNo ratings yet
- Biologie Thema 5 Boek 5b Deel 2Document8 pagesBiologie Thema 5 Boek 5b Deel 2Dima NajiNo ratings yet
- Examen Biologie JuniDocument18 pagesExamen Biologie JuniIsmaïl BerdaïNo ratings yet
- Het ZENUWSTELSEL 3Document39 pagesHet ZENUWSTELSEL 3angeliqueboekelman100% (2)
- Hoofdstuk 7Document14 pagesHoofdstuk 7Navepreet KaurNo ratings yet
- SpiereDocument6 pagesSpiereLilithNo ratings yet
- Humane Embryogenesis 2Document9 pagesHumane Embryogenesis 2ViktoriaNo ratings yet
- TUTM Week 2Document9 pagesTUTM Week 2Jelte FolkertsmaNo ratings yet
- FulltextDocument111 pagesFulltextJoepKluwenNo ratings yet
- 1B1 Week 1 HC4 Ionkanalen en HartpotentialenDocument10 pages1B1 Week 1 HC4 Ionkanalen en HartpotentialenVera VoogtNo ratings yet
- UntitledDocument2 pagesUntitledMichiel KoolenNo ratings yet
- Anatomie Hersenen PDFDocument3 pagesAnatomie Hersenen PDFJulia WindhorstNo ratings yet
- Brein Samenvatting PDFDocument50 pagesBrein Samenvatting PDFAmy SavageNo ratings yet
- Piramide BanenDocument5 pagesPiramide BanendiederikNo ratings yet
- Neuroanatomie 1Document127 pagesNeuroanatomie 1Tamar FoppenNo ratings yet
- Ar Thro LogieDocument23 pagesAr Thro LogielieselvandermaesNo ratings yet
- Deel 3 Hoofdstukken 1-3Document20 pagesDeel 3 Hoofdstukken 1-3Bram GeubbelmansNo ratings yet
- An atDocument7 pagesAn atGosse KrikkeNo ratings yet
- Leerdoelen Taak 2Document8 pagesLeerdoelen Taak 2Bram WerneNo ratings yet
- Anatomie NederlandDocument67 pagesAnatomie NederlandTom StevensNo ratings yet
- Tut MDocument8 pagesTut MRaoul van der PloegNo ratings yet
- Biologie Samenvatting Hoofdstuk 1Document4 pagesBiologie Samenvatting Hoofdstuk 1Isa GroenenNo ratings yet
- Biologie SamenvattingDocument5 pagesBiologie SamenvattingValeria MolNo ratings yet
- Orgaansystemen PDFDocument48 pagesOrgaansystemen PDFMichiel HentenaarNo ratings yet
- Antwoorden InstaptoetsenDocument3 pagesAntwoorden InstaptoetsenLevi WNo ratings yet
- AtrokinematiecaDocument3 pagesAtrokinematiecaSabine Van T VeldNo ratings yet
- Belangrijk MSA KTDocument15 pagesBelangrijk MSA KTTessa HodesNo ratings yet
- SV Waarneming VWO2 Biologie Voor JouDocument8 pagesSV Waarneming VWO2 Biologie Voor JoujuliaNo ratings yet
- Embryologie Samenvatting HC 1-5Document14 pagesEmbryologie Samenvatting HC 1-5Robert-Jan ArkeNo ratings yet
- Samenvatting StatistiekDocument6 pagesSamenvatting StatistiekFloor VerbeekNo ratings yet
- Samenvatting Interne Geneeskunde Hoofdstuk 22Document25 pagesSamenvatting Interne Geneeskunde Hoofdstuk 22Floor VerbeekNo ratings yet
- Spierstelsel Onderbeen en VoetDocument31 pagesSpierstelsel Onderbeen en VoetFloor VerbeekNo ratings yet
- Fysiologie Maag-DarmKanaalDocument31 pagesFysiologie Maag-DarmKanaalFloor VerbeekNo ratings yet
- Histologie en Celbiologie MDLDocument19 pagesHistologie en Celbiologie MDLFloor VerbeekNo ratings yet
- Basisbegrippen FarmacologieDocument10 pagesBasisbegrippen FarmacologieFloor VerbeekNo ratings yet
- Anatomie Van Het BovenbeenDocument13 pagesAnatomie Van Het BovenbeenFloor VerbeekNo ratings yet
- Anatomie Van de HeupDocument13 pagesAnatomie Van de HeupFloor VerbeekNo ratings yet