Download as pdf or txt
You might also like
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5820)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1093)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (845)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (590)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (898)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (540)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (349)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (822)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (122)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (401)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (74)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- Iodine - The Universal and Holistic Super Mineral - Dr. Gabriel CousensDocument32 pagesIodine - The Universal and Holistic Super Mineral - Dr. Gabriel CousensJ.J.100% (7)
- January 2015 (IAL) QP - Unit 1 Edexcel Biology A-LevelDocument25 pagesJanuary 2015 (IAL) QP - Unit 1 Edexcel Biology A-LevelAyse KerimNo ratings yet
- Brainwave - Cymatic Frequency ListingDocument26 pagesBrainwave - Cymatic Frequency Listingronan2890No ratings yet
- A Detailed Lesson Plan in Science 10 (The Endocrine System)Document8 pagesA Detailed Lesson Plan in Science 10 (The Endocrine System)Renz CruzNo ratings yet
- Blood Sugar Reg-WPS OfficeDocument8 pagesBlood Sugar Reg-WPS Officetanjidul islamNo ratings yet
- G9 Summative TestDocument4 pagesG9 Summative Testmarisa canoNo ratings yet
- PBL 1 (Pneumonia)Document11 pagesPBL 1 (Pneumonia)Rakeesh VeeraNo ratings yet
- Cell Organelles Worksheet-PrintDocument2 pagesCell Organelles Worksheet-PrintmuraliMu0% (1)
- Workbook ActivityDocument2 pagesWorkbook ActivityRisciella 18No ratings yet
- 11 Revised Biology 21 PDFDocument10 pages11 Revised Biology 21 PDFShubhendu ChattopadhyayNo ratings yet
- Neuropathic Pain: Vanja Basic-Kes/ Iris Zavoreo/ Marijana Bosnar Mira Ivankovic", Milan Bitunjac'Document7 pagesNeuropathic Pain: Vanja Basic-Kes/ Iris Zavoreo/ Marijana Bosnar Mira Ivankovic", Milan Bitunjac'Meidy ShadanaNo ratings yet
- Postul Intermitent (Intermittent Fasting) Pentru SlabireDocument3 pagesPostul Intermitent (Intermittent Fasting) Pentru SlabireSeb SaabNo ratings yet
- TheratogsDocument19 pagesTheratogssamurai_sanju6026No ratings yet
- Deep Metaproteomic Analysis of Human Salivary SupernatantDocument18 pagesDeep Metaproteomic Analysis of Human Salivary SupernatantJobin John PR15BI1001No ratings yet
- Cholorophyll Content in Five Different Species of PlantDocument14 pagesCholorophyll Content in Five Different Species of Plantlatest tamil moviesNo ratings yet
- Ethanol Tolerance in Bacteria PDFDocument2 pagesEthanol Tolerance in Bacteria PDFReneeNo ratings yet
- Abs Bench 500 Manual 3esDocument15 pagesAbs Bench 500 Manual 3esmaremotusNo ratings yet
- Microbial Genetics: Julia Hartati, DRDocument21 pagesMicrobial Genetics: Julia Hartati, DRerwan_mivecNo ratings yet
- Pullups Workout ProgramDocument6 pagesPullups Workout ProgramZe Luis TerrassoNo ratings yet
- RBCs - Abnormal - Morphology FinalDocument33 pagesRBCs - Abnormal - Morphology FinalInahkoni Alpheus Sky OiragasNo ratings yet
- Biochem 34 FR Expt #7 & 9Document10 pagesBiochem 34 FR Expt #7 & 9louize_1496No ratings yet
- Regulasi CA, MG and PO4Document16 pagesRegulasi CA, MG and PO4siskaNo ratings yet
- S TPHA Liquid: Design VerificationDocument9 pagesS TPHA Liquid: Design VerificationTanveerNo ratings yet
- AP Psychology Forest Grove High School Mr. TusowDocument81 pagesAP Psychology Forest Grove High School Mr. TusowIsabella TNo ratings yet
- Bacterial Growth CurveDocument3 pagesBacterial Growth CurveGampa Vijaykumar100% (1)
- Soal Kidney CompiledDocument28 pagesSoal Kidney Compiledstella pangestika100% (1)
- QowdoDocument1 pageQowdoRSCERIA KANDANGANNo ratings yet
- Answer All The QuestionsDocument3 pagesAnswer All The Questionsmaimonsabi74100% (1)
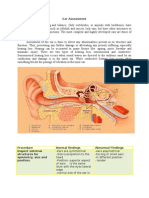
- Ear AssessmentDocument5 pagesEar AssessmentJake Amor100% (4)
- Calvin CycleDocument3 pagesCalvin Cyclekumarscribd5482No ratings yet