Download as pdf or txt
You might also like
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5820)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1093)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (845)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (590)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (898)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (540)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (349)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (822)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (122)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (401)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (74)
- CHC Exam HandbookDocument52 pagesCHC Exam Handbookkumar100% (3)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- 2 Rev CHT Guidebook 102411Document90 pages2 Rev CHT Guidebook 102411Sangkula Laja50% (2)
- Pedia With LeukemiaDocument14 pagesPedia With LeukemiaAlynna ValbuenaNo ratings yet
- Soluciones EjerciciosDocument10 pagesSoluciones EjerciciosJe RivasNo ratings yet
- Effects of Germanium Dioxide, An Inhibitor of Diatom Growth, On The Microscopic Laboratory Cultivation Stage of The Kelp, Laminaria SaccharinaDocument6 pagesEffects of Germanium Dioxide, An Inhibitor of Diatom Growth, On The Microscopic Laboratory Cultivation Stage of The Kelp, Laminaria SaccharinaJe RivasNo ratings yet
- Datos CanadaDocument14 pagesDatos CanadaJe RivasNo ratings yet
- 4 3 23 246Document6 pages4 3 23 246Je RivasNo ratings yet
- GC 2010ErrMsgDocument18 pagesGC 2010ErrMsgJe RivasNo ratings yet
- Accepted Manuscript: 10.1016/j.aca.2016.05.014Document34 pagesAccepted Manuscript: 10.1016/j.aca.2016.05.014Je RivasNo ratings yet
- Biological Conservation: Anna Kuczyn Ska, Evelyn MoorkensDocument8 pagesBiological Conservation: Anna Kuczyn Ska, Evelyn MoorkensJe RivasNo ratings yet
- Policosanol Content and Composition in Perilla Seeds: P A, K T H, J N P, C K KDocument4 pagesPolicosanol Content and Composition in Perilla Seeds: P A, K T H, J N P, C K KJe RivasNo ratings yet
- Fundamentals of Dairy Chemistry 3rd Ed - Noble P. Wong (Aspen Publishers, Inc. 1999) PDFDocument734 pagesFundamentals of Dairy Chemistry 3rd Ed - Noble P. Wong (Aspen Publishers, Inc. 1999) PDFJe RivasNo ratings yet
- Isolation, Identification and Characterization of Effective Bacteria On Bioremediation From The Waste Parts of Stevia Rebaudiana BerutoniDocument7 pagesIsolation, Identification and Characterization of Effective Bacteria On Bioremediation From The Waste Parts of Stevia Rebaudiana BerutoniJe RivasNo ratings yet
- Analytical Biochemistry: Recombinant Expression of Aryl Hydrocarbon Receptor For Quantitative Ligand-Binding AnalysisDocument9 pagesAnalytical Biochemistry: Recombinant Expression of Aryl Hydrocarbon Receptor For Quantitative Ligand-Binding AnalysisJe RivasNo ratings yet
- Ace TilaDocument4 pagesAce TilaJe RivasNo ratings yet
- Evaluating Microtox As A Tool For Biodegradability Assessment of Partially Treated Solutions of Pesticides Using Fe and Tio Solar Photo-Assisted ProcessesDocument10 pagesEvaluating Microtox As A Tool For Biodegradability Assessment of Partially Treated Solutions of Pesticides Using Fe and Tio Solar Photo-Assisted ProcessesJe RivasNo ratings yet
- Journal of Chromatography A, 1218 (2011) 8480-8489Document10 pagesJournal of Chromatography A, 1218 (2011) 8480-8489Je RivasNo ratings yet
- The in Uence of Soil Heavy Metals Pollution On Soil Microbial Biomass, Enzyme Activity, and Community Composition Near A Copper SmelterDocument7 pagesThe in Uence of Soil Heavy Metals Pollution On Soil Microbial Biomass, Enzyme Activity, and Community Composition Near A Copper SmelterJe RivasNo ratings yet
- Forensic Science International 142 (2004) 161-210Document50 pagesForensic Science International 142 (2004) 161-210Je RivasNo ratings yet
- Colloids and Surfaces A. Physicochem. Eng. Aspects 360 (2010) 6-12Document7 pagesColloids and Surfaces A. Physicochem. Eng. Aspects 360 (2010) 6-12Je RivasNo ratings yet
- EUA Ihealth RapidTestAg ifuHT - 0 - 0Document14 pagesEUA Ihealth RapidTestAg ifuHT - 0 - 0thehumansyndromeNo ratings yet
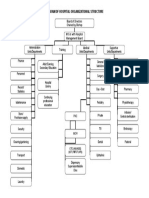
- Diagram of Hospital Organizational Structure: Board of Directors Chaired by Bishop MO I/c With Hospital Management BoardDocument1 pageDiagram of Hospital Organizational Structure: Board of Directors Chaired by Bishop MO I/c With Hospital Management BoardBELAY AWOKENo ratings yet
- Lancaster, John E. - Newcastle Disease - A ReviDocument201 pagesLancaster, John E. - Newcastle Disease - A RevipaigneilNo ratings yet
- Mineire'sDocument16 pagesMineire'sShaykh Evangelista BacualNo ratings yet
- Tomato Foliage ProblemsDocument7 pagesTomato Foliage ProblemsFandi Ahmad SNo ratings yet
- Aminoglycosides: Aminoglycoside Is CategoryDocument6 pagesAminoglycosides: Aminoglycoside Is CategoryAnonymous RJwbBCkrHNo ratings yet
- Clarito LDocument3 pagesClarito LMaria Danice AngelaNo ratings yet
- 1000 Mcqs - MiscellaneousDocument11 pages1000 Mcqs - MiscellaneousSelvaArockiamNo ratings yet
- The Importance of Vaccination During A PandemicDocument2 pagesThe Importance of Vaccination During A PandemicSalsabilla KusumaningrumNo ratings yet
- Both Young and Old RBC Young RBC Young RBC Old RBC: Rizal ST., Albay Dist., Legazpi CityDocument3 pagesBoth Young and Old RBC Young RBC Young RBC Old RBC: Rizal ST., Albay Dist., Legazpi CityAbegail ListancoNo ratings yet
- Pathology Question BankDocument65 pagesPathology Question BankJIBIN JOSE100% (1)
- HIV in South AfricaDocument17 pagesHIV in South AfricaMadelineNo ratings yet
- EBOLADocument2 pagesEBOLAamirNo ratings yet
- Parasitology: Course 1Document57 pagesParasitology: Course 1anaNo ratings yet
- Laporan-Analisa-SKDR (1)Document3 pagesLaporan-Analisa-SKDR (1)Puskesmas LampasiNo ratings yet
- Schisto. in The Phils. & CaragaDocument56 pagesSchisto. in The Phils. & CaragaHoneylet Ü Ferol100% (2)
- PMK No. 49 Tahun 2013 TTG Komite Keperawatan RS-capDocument66 pagesPMK No. 49 Tahun 2013 TTG Komite Keperawatan RS-capyullies ekaNo ratings yet
- GeneXpert SOP Xpert Processing EPTB Specimens DRAFTDocument7 pagesGeneXpert SOP Xpert Processing EPTB Specimens DRAFTFitra lestari100% (1)
- DafpusDocument3 pagesDafpusNurizza RahmaniaNo ratings yet
- PneumoniaDocument10 pagesPneumoniaDina M ElbakaryNo ratings yet
- White Blood CellsDocument2 pagesWhite Blood Cellsهاشم ابوطالبNo ratings yet
- Rangkuman Week 5 (PBL) - OsteomyelitisDocument9 pagesRangkuman Week 5 (PBL) - OsteomyelitisMaria Dini AdmiratiNo ratings yet
- Deep Clearing April 231Document5 pagesDeep Clearing April 231Javier V100% (2)
- Safe Injection and Infusion PracticesDocument2 pagesSafe Injection and Infusion PracticesholyfamilyNo ratings yet
- Fenotipovi Astme Kod DjeceDocument6 pagesFenotipovi Astme Kod Djecezijad_lNo ratings yet
- Masro - Protozoa Helminth AntropodsDocument34 pagesMasro - Protozoa Helminth AntropodsAnis AqilahNo ratings yet
- Canine - Leptospirosis Marge PoughDocument1 pageCanine - Leptospirosis Marge Poughw00fNo ratings yet