Download as pdf or txt
You might also like
- Struttura Uomo - enDocument304 pagesStruttura Uomo - enNekro TopsNo ratings yet
- Kerala Syllabus BamsDocument70 pagesKerala Syllabus BamsSwastik SureshNo ratings yet
- Wound Documentation TipsDocument4 pagesWound Documentation TipsLaura Hernandez100% (1)
- Nornltates: A Review of The Chilean Spiders of The Superfamily Migoidea (Araneae, Mygalomorphae)Document16 pagesNornltates: A Review of The Chilean Spiders of The Superfamily Migoidea (Araneae, Mygalomorphae)Lucas MedeirosNo ratings yet
- A New Species of BathymedonDocument16 pagesA New Species of BathymedonraibpNo ratings yet
- Handbook SpintheridaeDocument7 pagesHandbook Spintheridaeneimar5003No ratings yet
- Budiewi Et Al 2020Document10 pagesBudiewi Et Al 2020heimat66No ratings yet
- Ournal of Atural IstoryDocument20 pagesOurnal of Atural IstoryANALIZEEEEEEENo ratings yet
- A New ?lamellipedian Arthropod From The Early Cambrian Sirius Passet Fauna of North GreenlandDocument7 pagesA New ?lamellipedian Arthropod From The Early Cambrian Sirius Passet Fauna of North GreenlandSukma MaulidaNo ratings yet
- Martin-Park Et Al. (2012) - Revision of Genus Repipta Stal 1859Document54 pagesMartin-Park Et Al. (2012) - Revision of Genus Repipta Stal 1859caritocastri1986No ratings yet
- Redescription of Female Laelaps NuttalliDocument9 pagesRedescription of Female Laelaps Nuttallimelanieruiz09No ratings yet
- Carlos Lorenzo & Juan Luis ArsuagaDocument22 pagesCarlos Lorenzo & Juan Luis ArsuagatrutiNo ratings yet
- Taxonomic Study of The Family Scoliidae (Hymenoptera Aculeata) in IraqDocument15 pagesTaxonomic Study of The Family Scoliidae (Hymenoptera Aculeata) in IraqInternational Network For Natural SciencesNo ratings yet
- Andia & Grismado - RevMACN - 17-1 - PaginadoDocument6 pagesAndia & Grismado - RevMACN - 17-1 - Paginadoaliciacardozo63No ratings yet
- Katayama Et Al. - 2012 - A New Species of Trichonotus From SomaliaDocument13 pagesKatayama Et Al. - 2012 - A New Species of Trichonotus From SomaliaMuntasir AkashNo ratings yet
- Bela MinoicaDocument6 pagesBela MinoicaRiccardo Giannuzzi SavelliNo ratings yet
- Cephalic Sensorial Pores in Galaxiidae From Chile (Saez y Pequeño, 2014)Document6 pagesCephalic Sensorial Pores in Galaxiidae From Chile (Saez y Pequeño, 2014)Francisco J. OvalleNo ratings yet
- Larval A. Simplex BDocument8 pagesLarval A. Simplex Bape77No ratings yet
- Enigmatic Amphibians in Mid-Cretaceous Amber Were Chameleon-Like Ballistic FeedersDocument6 pagesEnigmatic Amphibians in Mid-Cretaceous Amber Were Chameleon-Like Ballistic FeedersDirtyBut WholeNo ratings yet
- A New Fossil Genus Near Meghyperus Loew From Baltic Amber (Diptera: Atelestidae)Document4 pagesA New Fossil Genus Near Meghyperus Loew From Baltic Amber (Diptera: Atelestidae)agnieszka.soszynska.majNo ratings yet
- 3 2002232Ng241360Document21 pages3 2002232Ng241360Juba chabaneNo ratings yet
- Clark (1970) Loss of The Left Oviduct in The Colubrid Snake Genus TantillaDocument5 pagesClark (1970) Loss of The Left Oviduct in The Colubrid Snake Genus TantillaDiego LoboNo ratings yet
- 2007 Oreobates Lehri Padial EtalDocument9 pages2007 Oreobates Lehri Padial EtalJuanca Chaparro AuzaNo ratings yet
- Funtional Morphology of Bite Mechanics in The Great BarracudaDocument14 pagesFuntional Morphology of Bite Mechanics in The Great BarracudaassyahwaNo ratings yet
- Aesa/58 2 247Document2 pagesAesa/58 2 247Juan Enrique Tacoronte MoralesNo ratings yet
- Formas.1976. Descriptions of Batrachyla TadpolesDocument6 pagesFormas.1976. Descriptions of Batrachyla TadpolesTerreno CicloFaunaNo ratings yet
- Punya Ata Petir 2Document10 pagesPunya Ata Petir 2Rian Dani TumanggorNo ratings yet
- Boldrini Et Al., 2017 (Brasil) BaDocument10 pagesBoldrini Et Al., 2017 (Brasil) Bacarmen villalobosNo ratings yet
- Baqri&Coomans 1973 - Dorylaimida Descritos Por Stekhoven y Teunissen 1938Document57 pagesBaqri&Coomans 1973 - Dorylaimida Descritos Por Stekhoven y Teunissen 1938Miguelillo_HernandezNo ratings yet
- 79989-Article Text-291227-319115-10-20230104Document9 pages79989-Article Text-291227-319115-10-20230104sharktank4youNo ratings yet
- Hero 1989 Code For Toe Clipping AnuransDocument2 pagesHero 1989 Code For Toe Clipping Anuranslaspiur22blues7327No ratings yet
- 10anadara Subrubra Dunker 1866Document11 pages10anadara Subrubra Dunker 1866Mhd FaroukNo ratings yet
- 25718-Article Text-70217-83062-10-20161019Document18 pages25718-Article Text-70217-83062-10-20161019NatanAntonioliNo ratings yet
- Avicularia Rickwesti Sp. Nov., A Remarkable New Species of AviculariaDocument5 pagesAvicularia Rickwesti Sp. Nov., A Remarkable New Species of AviculariazzzaNo ratings yet
- Kostka 1991Document12 pagesKostka 1991Lola BoaNo ratings yet
- A New Genus and Species of Grass SpecialDocument10 pagesA New Genus and Species of Grass SpecialElidiomar Ribeiro da SilvaNo ratings yet
- Doyle Et Al. (1990) - L-J To E-K Macrofossils From Weddel Sea, AntarcticaDocument6 pagesDoyle Et Al. (1990) - L-J To E-K Macrofossils From Weddel Sea, AntarcticaArturo Palma RamírezNo ratings yet
- 11 Sandoval LeivaDocument3 pages11 Sandoval LeivaPaula AlisterNo ratings yet
- 2009 Zool 26 (1) 155-160Document6 pages2009 Zool 26 (1) 155-160Katherine Llerena CalderónNo ratings yet
- Zeugopterus RegiusDocument3 pagesZeugopterus Regiusantonpc100% (2)
- Mantophasmatodea: A New Insect Order With Extant Members in The AfrotropicsDocument5 pagesMantophasmatodea: A New Insect Order With Extant Members in The AfrotropicsagccNo ratings yet
- COPA4348 ReprintDocument7 pagesCOPA4348 ReprintaquarogioNo ratings yet
- Marshal and Thornton 1963. MICROMALTHUS (Coleoptera - Micromalthidae) in Hong KongDocument6 pagesMarshal and Thornton 1963. MICROMALTHUS (Coleoptera - Micromalthidae) in Hong KongCarlos AguilarNo ratings yet
- New Record of CaprellaDocument6 pagesNew Record of CaprellaAna VerdiNo ratings yet
- A New Species of Parameiropsis Becker 1974 CopepodDocument12 pagesA New Species of Parameiropsis Becker 1974 CopepodpcorgoNo ratings yet
- Ma 2006 J Euk Micro Epistylis Onblue CrabDocument11 pagesMa 2006 J Euk Micro Epistylis Onblue CrabRIZKI NANDANo ratings yet
- Bray 1996Document13 pagesBray 1996Luis angelNo ratings yet
- Howden Gill 1995Document7 pagesHowden Gill 1995Maryzender Rodríguez MelgarejoNo ratings yet
- Neotropical Acroceridae KeysDocument36 pagesNeotropical Acroceridae KeysPedroVaNo ratings yet
- Unusual Teratological Cases in Scarabaeidae (Coleoptera: Scarabaeoidea) : Two Specimens With Multiple MalformationsDocument7 pagesUnusual Teratological Cases in Scarabaeidae (Coleoptera: Scarabaeoidea) : Two Specimens With Multiple Malformationsleonello milaniNo ratings yet
- New Combinations in Thai FernsDocument7 pagesNew Combinations in Thai FernsruddhasenNo ratings yet
- Kim y Jung 2018Document6 pagesKim y Jung 2018Antonela HerreraNo ratings yet
- A New Species of Burrowing CrayfishDocument13 pagesA New Species of Burrowing CrayfishDavid Orlando Carmona LagosNo ratings yet
- Zaragoza Tapia 2020aDocument16 pagesZaragoza Tapia 2020aWilliam Scott Monks SheetsNo ratings yet
- Khan & Asfaq 2005aDocument4 pagesKhan & Asfaq 2005aabdulraheemabdullah56No ratings yet
- New Records On Hermit CrabsDocument8 pagesNew Records On Hermit CrabsNuzaiba P muhammedNo ratings yet
- 2002 Oreailurus Jacobita Description Comparision Felines Altiplano - GarciaPereaDocument15 pages2002 Oreailurus Jacobita Description Comparision Felines Altiplano - GarciaPereaLucía HNo ratings yet
- Zootaxa: CorrespondenceDocument5 pagesZootaxa: Correspondencecabruca1No ratings yet
- 17 - Polhemus Spangler 1995Document25 pages17 - Polhemus Spangler 1995oséias martins magalhãesNo ratings yet
- Halictophagidae AustraliaDocument39 pagesHalictophagidae AustraliaKania TrieswandaNo ratings yet
- Faraji 2006Document4 pagesFaraji 2006abdulraheemabdullah56No ratings yet
- Celestial Pearl Danio Is A Miniature DanioDocument28 pagesCelestial Pearl Danio Is A Miniature DaniopccostiNo ratings yet
- Systematic Status of a South American Frog, Allophryne ruthveni GaigeFrom EverandSystematic Status of a South American Frog, Allophryne ruthveni GaigeNo ratings yet
- Komrkov 2015Document10 pagesKomrkov 2015Hernan Turpo AroquipaNo ratings yet
- Harris 2016Document9 pagesHarris 2016Hernan Turpo AroquipaNo ratings yet
- Bertani 2017Document7 pagesBertani 2017Hernan Turpo AroquipaNo ratings yet
- Science of The Total EnvironmentDocument15 pagesScience of The Total EnvironmentHernan Turpo AroquipaNo ratings yet
- Effects of Environmental Factors On The Uptake Rates of Dissolved Nitrogen by A Salt-Water Green Alga (Oocystis Borgei Snow)Document5 pagesEffects of Environmental Factors On The Uptake Rates of Dissolved Nitrogen by A Salt-Water Green Alga (Oocystis Borgei Snow)Hernan Turpo AroquipaNo ratings yet
- Berto Lo 2010Document8 pagesBerto Lo 2010Hernan Turpo AroquipaNo ratings yet
- Copepoda: Order - CalanoidaDocument10 pagesCopepoda: Order - CalanoidaHernan Turpo Aroquipa100% (1)
- Preliminary Zooplankton Study of Six "Mercedinas" Lakes (San Luis, Argentina)Document8 pagesPreliminary Zooplankton Study of Six "Mercedinas" Lakes (San Luis, Argentina)Hernan Turpo AroquipaNo ratings yet
- A Revision of Infragenerics Taxa of FissidensDocument21 pagesA Revision of Infragenerics Taxa of FissidensCristopher Jimenez OrozcoNo ratings yet
- Shiatsu MassageDocument13 pagesShiatsu MassageLawrence Cada Nofies100% (1)
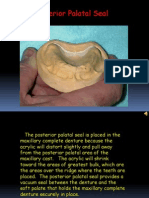
- Making A Posterior Palatal SealDocument16 pagesMaking A Posterior Palatal SealmegamarwaNo ratings yet
- Laquan McDonald Police Reports, Part 3Document60 pagesLaquan McDonald Police Reports, Part 3jroneillNo ratings yet
- Chapter 1: The Human Body: An Orientation: Anatomy - The Study of StructureDocument7 pagesChapter 1: The Human Body: An Orientation: Anatomy - The Study of StructureptldhrbNo ratings yet
- Buccal Fat Pad Flap-1Document4 pagesBuccal Fat Pad Flap-1Alex OprisanNo ratings yet
- Surugiu (2023) A New Species of Scolelepis From The Black SeaDocument21 pagesSurugiu (2023) A New Species of Scolelepis From The Black SeaVictor SurugiuNo ratings yet
- Chest PhysiotherapyDocument22 pagesChest PhysiotherapyGhia_Angeles_8457No ratings yet
- 18 - Elbow TraumaDocument42 pages18 - Elbow Traumadrcoolcat2000No ratings yet
- Third Division G.R. No. 229829, January 22, 2018 PEOPLE OF THE PHILIPPINES, Plaintiff-Appellee, v. ARNEL KALIPAYAN Y ANIANO, Accused-Appellant. Decision Gesmundo, J.Document20 pagesThird Division G.R. No. 229829, January 22, 2018 PEOPLE OF THE PHILIPPINES, Plaintiff-Appellee, v. ARNEL KALIPAYAN Y ANIANO, Accused-Appellant. Decision Gesmundo, J.Reymar BurceNo ratings yet
- Study of The Morphometry and Meristic Analyses of Three Mystus Species From The Chandubi Beel, Kamrup District, AssamDocument10 pagesStudy of The Morphometry and Meristic Analyses of Three Mystus Species From The Chandubi Beel, Kamrup District, AssamSSR-IIJLS JournalNo ratings yet
- Yoga Teacher Training Anatomy of Movement For Yoga TeachersDocument22 pagesYoga Teacher Training Anatomy of Movement For Yoga TeachersMarkusNo ratings yet
- Suspensory Ligament Rupture Tech During Small Animal OH PDFDocument8 pagesSuspensory Ligament Rupture Tech During Small Animal OH PDFRiesky NudialestariNo ratings yet
- Radposi Lesson 1 and 2Document17 pagesRadposi Lesson 1 and 2Cesarina BlancaNo ratings yet
- Hassel 1995Document9 pagesHassel 1995Emilio MurciaNo ratings yet
- Early Silurian Trilobites AustraliaDocument41 pagesEarly Silurian Trilobites AustraliaardeegeeNo ratings yet
- Dieseldorf Collection: Ceramic Corpus of The Maya Terminal Classic Originating From MoldsDocument48 pagesDieseldorf Collection: Ceramic Corpus of The Maya Terminal Classic Originating From MoldsPablo AlarcónNo ratings yet
- Lecture 04 PDFDocument9 pagesLecture 04 PDFNuta Marius AlexandruNo ratings yet
- PPTDocument17 pagesPPTnimi mem0nNo ratings yet
- Teaching Chinese Pulse ImagesDocument7 pagesTeaching Chinese Pulse ImagesKeefer Michael100% (1)
- Chen BingDocument26 pagesChen Bingsuagde1302No ratings yet
- The Topography of Upper LimbDocument5 pagesThe Topography of Upper LimbLorena DobrescuNo ratings yet
- Thesis by Woong Kim SmallerDocument415 pagesThesis by Woong Kim SmallerWoong KimNo ratings yet
- Current Topics On Fetal 3D 4D Ultrasound 2018Document256 pagesCurrent Topics On Fetal 3D 4D Ultrasound 2018DRDRAGON2008100% (1)
- Criminal Law Cases For Better Understanding of Art 14Document392 pagesCriminal Law Cases For Better Understanding of Art 14AweGooseTreeNo ratings yet
- ANAT 100 - Module 1 Summary NotesDocument6 pagesANAT 100 - Module 1 Summary NotesRahul BhardwajNo ratings yet
- Earthworm BehaviorsDocument6 pagesEarthworm BehaviorskellymcnameeNo ratings yet