Download as doc, pdf, or txt
You might also like
- Miller Anesthesia 7th Ed Chapter 34 Preop EvaluationDocument66 pagesMiller Anesthesia 7th Ed Chapter 34 Preop Evaluationjbahalkeh100% (1)
- 10 Tips For A Better BackDocument18 pages10 Tips For A Better BackPhysio Ebook100% (1)
- Update of Survey, Regulation and Toxic Effects of Mycotoxins in EuropeDocument10 pagesUpdate of Survey, Regulation and Toxic Effects of Mycotoxins in EuropeHigor MarkovicNo ratings yet
- SummaryefsadonDocument1 pageSummaryefsadonIGFAieNo ratings yet
- 2022 - Variation in Tocochromanols Level and Mycotoxins Content in Sweet MaizeDocument13 pages2022 - Variation in Tocochromanols Level and Mycotoxins Content in Sweet Maizepawel.gornasNo ratings yet
- KSU Mycotoxins in Swine Diets Fact SheetDocument6 pagesKSU Mycotoxins in Swine Diets Fact SheetEva Estel García VelásquezNo ratings yet
- Toxins 16 00154Document14 pagesToxins 16 00154aqalvaro6471No ratings yet
- Toxins: Microbial Inhibition of Fusarium Pathogens and Biological Modification of Trichothecenes in Cereal GrainsDocument14 pagesToxins: Microbial Inhibition of Fusarium Pathogens and Biological Modification of Trichothecenes in Cereal GrainsFernanda SilvaNo ratings yet
- Antifungal PDFDocument34 pagesAntifungal PDFUmashankar SharmaNo ratings yet
- Eco-Friendly Bacterial Biodegradation of Mycotoxins: Review ArticleDocument10 pagesEco-Friendly Bacterial Biodegradation of Mycotoxins: Review ArticleSyed Ali Akbar BokhariNo ratings yet
- Comparative in Vitro Assessment of A Range of Commercial Feed Additives With Multiple Mycotoxin Binding ClaimsDocument14 pagesComparative in Vitro Assessment of A Range of Commercial Feed Additives With Multiple Mycotoxin Binding ClaimsSumankeerthiNo ratings yet
- Mycotoxicosis and MycetismusDocument31 pagesMycotoxicosis and MycetismusLincy JohnyNo ratings yet
- Bacteriology v7 Id1130Document3 pagesBacteriology v7 Id1130AntaraaNo ratings yet
- The Pulmonary Mycoses: Aaron Samuel Miller, MD, MSPH, and Robert William Wilmott, BSC, MB, BS, MD, FRCP (Uk)Document25 pagesThe Pulmonary Mycoses: Aaron Samuel Miller, MD, MSPH, and Robert William Wilmott, BSC, MB, BS, MD, FRCP (Uk)Sarah Fitriani MuzwarNo ratings yet
- The Pulmonary Mycoses: Aaron Samuel Miller, MD, MSPH, and Robert William Wilmott, BSC, MB, BS, MD, FRCP (Uk)Document25 pagesThe Pulmonary Mycoses: Aaron Samuel Miller, MD, MSPH, and Robert William Wilmott, BSC, MB, BS, MD, FRCP (Uk)Klinikdr RIDHANo ratings yet
- Baysal, Çalışkan, Yeşilova - 2008 - An Inhibitory Effect of A New Bacillus Subtilis Strain (EU07) Against Fusarium Oxysporum F. Sp. Radi PDFDocument8 pagesBaysal, Çalışkan, Yeşilova - 2008 - An Inhibitory Effect of A New Bacillus Subtilis Strain (EU07) Against Fusarium Oxysporum F. Sp. Radi PDFJosé Matías ZapiolaNo ratings yet
- Rajiv Gandhi Institute of Veterinary Education and Research: "Mycotoxins in Duck"Document7 pagesRajiv Gandhi Institute of Veterinary Education and Research: "Mycotoxins in Duck"Sree DharNo ratings yet
- MycotoxinDocument28 pagesMycotoxin064 Poonguzhali.SNo ratings yet
- Toxins 13 00148 v2Document22 pagesToxins 13 00148 v2Cristina BulgaruNo ratings yet
- Zebrafish AnesthesiaDocument23 pagesZebrafish AnesthesiaTania PereraNo ratings yet
- The Role of Environmental Exposures in Atopic DermatitisDocument18 pagesThe Role of Environmental Exposures in Atopic DermatitisRatih Anindita Rahajeng RipyonoNo ratings yet
- Toxins 15 00653Document17 pagesToxins 15 00653aqalvaro6471No ratings yet
- Aflaxoxin ToxicityDocument13 pagesAflaxoxin Toxicitycorneliusrono1995No ratings yet
- Toxins 16 00003Document10 pagesToxins 16 00003aqalvaro6471No ratings yet
- Ionophore ToxicityDocument15 pagesIonophore ToxicityKamini AnamaleiNo ratings yet
- Pseudomonas Fluorescens A Potential Bacterial Antagonist To Control Plant DiseasesDocument13 pagesPseudomonas Fluorescens A Potential Bacterial Antagonist To Control Plant DiseasesPalak AgarwalNo ratings yet
- Toxic Effects of Organochlorine Pesticides: A Review: American Journal of BioscienceDocument8 pagesToxic Effects of Organochlorine Pesticides: A Review: American Journal of BioscienceBalogun YusufNo ratings yet
- Impact of Malnutrition On Immunity and InfectionDocument17 pagesImpact of Malnutrition On Immunity and InfectionYusuf NugrohoNo ratings yet
- Tropicalmed 03 00037Document9 pagesTropicalmed 03 00037dianisaindiraNo ratings yet
- Reduced Tomato Bacterial Wilt by Ferrous ChlorideDocument6 pagesReduced Tomato Bacterial Wilt by Ferrous Chloridechamarawmas95No ratings yet
- Feed Additives To Control Poultry: Salmonella inDocument13 pagesFeed Additives To Control Poultry: Salmonella inelsa lestariNo ratings yet
- Aflatoxin in Food: Implications On Health and Control MeasuresDocument7 pagesAflatoxin in Food: Implications On Health and Control MeasuresVi-e -minNo ratings yet
- Mycotoxins On Poultry Production: AspergillusDocument12 pagesMycotoxins On Poultry Production: AspergillusmalwyliNo ratings yet
- Toxic Effects of Fluoride On OrganismsDocument7 pagesToxic Effects of Fluoride On OrganismsFranciele AnjosNo ratings yet
- 317567fumonisin B1Document3 pages317567fumonisin B1tophesnyksNo ratings yet
- MoleculesDocument13 pagesMoleculessplaingNo ratings yet
- Acute Toxoplasmosis and AntioxidantDocument7 pagesAcute Toxoplasmosis and AntioxidantVanessa VítorinoNo ratings yet
- Pseudomona Aeruginosa Long 2012Document7 pagesPseudomona Aeruginosa Long 2012Laura López Del Castillo LalydelcaNo ratings yet
- Biology 11 01602 v2Document17 pagesBiology 11 01602 v2mcbio2023.2No ratings yet
- Elevated Systemic Antibodies Towards Commensal Gut Microbiota in Autoinflammatory ConditionDocument10 pagesElevated Systemic Antibodies Towards Commensal Gut Microbiota in Autoinflammatory ConditionFrontiersNo ratings yet
- Management of Bacterial Wilt (Ralstonia Solanacearum) of Potato: Focus On Natural Bioactive CompoundsDocument20 pagesManagement of Bacterial Wilt (Ralstonia Solanacearum) of Potato: Focus On Natural Bioactive Compoundsdawit gNo ratings yet
- Noureenetal 2023Document7 pagesNoureenetal 2023seithshazamNo ratings yet
- MycotoxinsDocument4 pagesMycotoxinsnepoliyan ghimireNo ratings yet
- Chapter Ii-RrlDocument10 pagesChapter Ii-RrlmaeshengNo ratings yet
- Denagard Respiratory and Enteric Review (U. Klein)Document8 pagesDenagard Respiratory and Enteric Review (U. Klein)nick224No ratings yet
- Chem 121 PDFDocument12 pagesChem 121 PDFmasuma lovelyNo ratings yet
- Applied Soil EcologyDocument12 pagesApplied Soil EcologyEnry JimenezNo ratings yet
- Economic Effect Parasite Assignment 3Document4 pagesEconomic Effect Parasite Assignment 3MUHAMMADNo ratings yet
- Ssjcoe Manufacture of FluconazoleDocument83 pagesSsjcoe Manufacture of FluconazoleBhavik PanchalNo ratings yet
- Lectura Primer Previo 2021Document14 pagesLectura Primer Previo 2021JOSE GERARDO GONZALEZ YANEZNo ratings yet
- 3) The Role and Importance of Haccp: ParasitesDocument10 pages3) The Role and Importance of Haccp: ParasitesNirmal SharmaNo ratings yet
- Clostridium Botulinum Neurotoxins ProducedDocument5 pagesClostridium Botulinum Neurotoxins Producedcatsoul87No ratings yet
- Provided For Non-Commercial Research and Educational Use Only. Not For Reproduction, Distribution or Commercial UseDocument7 pagesProvided For Non-Commercial Research and Educational Use Only. Not For Reproduction, Distribution or Commercial Usemathan kumarNo ratings yet
- M7L7B1 The Endotoxin Hypothesis of NeurodegenerationDocument10 pagesM7L7B1 The Endotoxin Hypothesis of NeurodegenerationLeocadio Alcalá MORILLASNo ratings yet
- Aflatoxins: Aspergillus. Parasiticus. They Are Produce and Widely Distributed in The Tropical and SubtropicalDocument5 pagesAflatoxins: Aspergillus. Parasiticus. They Are Produce and Widely Distributed in The Tropical and SubtropicalebenezerNo ratings yet
- Pseudomonas Fluorescens: General CharacteristicsDocument4 pagesPseudomonas Fluorescens: General CharacteristicsPriya Sharma100% (1)
- Inhibition of Production of Cyclopiazonic Acid and Ochratoxin A by The Fungicide Iprodione+Document4 pagesInhibition of Production of Cyclopiazonic Acid and Ochratoxin A by The Fungicide Iprodione+DiegoSampietroNo ratings yet
- Application of Calcium and Sulfur in The Severity of Puccinia Coronata F. Sp. AvenaeDocument5 pagesApplication of Calcium and Sulfur in The Severity of Puccinia Coronata F. Sp. AvenaeIJAERS JOURNALNo ratings yet
- Immunotoxic Effects of Imidacloprid Following 28 Days of Oral Exposure in BALB/c MiceDocument11 pagesImmunotoxic Effects of Imidacloprid Following 28 Days of Oral Exposure in BALB/c MiceImam HasanNo ratings yet
- Control BiologicoDocument13 pagesControl BiologicomurielNo ratings yet
- Infectious Bursal Disease (GUMBORO Disease) in Chickens: Minalu Teshome, Tewodros Fentahunand Bemrew AdmassuDocument7 pagesInfectious Bursal Disease (GUMBORO Disease) in Chickens: Minalu Teshome, Tewodros Fentahunand Bemrew AdmassuDhaif dhaifNo ratings yet
- Insect ImmunologyFrom EverandInsect ImmunologyNancy E. BeckageNo ratings yet
- Raportarea Catre Oie A Unui Focar de BoalaDocument2 pagesRaportarea Catre Oie A Unui Focar de BoalaCotovici Catana AlinaNo ratings yet
- REFERAT - Salinomycin Intoxication in TurkeysDocument4 pagesREFERAT - Salinomycin Intoxication in TurkeysCotovici Catana AlinaNo ratings yet
- Compunere - New CafeteriaDocument1 pageCompunere - New CafeteriaCotovici Catana AlinaNo ratings yet
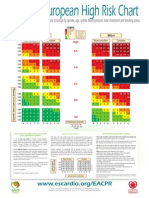
- 10 Year Risk of Fatal CVD in High Risk Regions of Europe by Gender, Age, Systolic Blood Pressure, Total Cholesterol and Smoking StatusDocument2 pages10 Year Risk of Fatal CVD in High Risk Regions of Europe by Gender, Age, Systolic Blood Pressure, Total Cholesterol and Smoking StatusCotovici Catana AlinaNo ratings yet
- Indications: PeritonitisDocument8 pagesIndications: PeritonitisIrene Soriano BayubayNo ratings yet
- Endoscopic Anatomy of Nose and PNS FinalDocument98 pagesEndoscopic Anatomy of Nose and PNS FinalRahul Kurkure100% (2)
- Heart FailureDocument43 pagesHeart FailuresimonjosanNo ratings yet
- 8BreathingExercises Calmhappyhealthy PDFDocument10 pages8BreathingExercises Calmhappyhealthy PDFlunn100% (1)
- Arman-2016-Stress Experience and Effect On Self-Perceived Oral Health Status Among High School StudentsDocument5 pagesArman-2016-Stress Experience and Effect On Self-Perceived Oral Health Status Among High School StudentsJontavius GilbertNo ratings yet
- Drugs For The Treatment of Diabetes MellitusDocument54 pagesDrugs For The Treatment of Diabetes MellitusAndreas AndreouNo ratings yet
- Pedia BADocument72 pagesPedia BABabbie CasinNo ratings yet
- Histology: First Semester A.Y. 2021 - 2022Document6 pagesHistology: First Semester A.Y. 2021 - 2022Fea Kristine PacquiaoNo ratings yet
- Deep Metaproteomic Analysis of Human Salivary SupernatantDocument18 pagesDeep Metaproteomic Analysis of Human Salivary SupernatantJobin John PR15BI1001No ratings yet
- Block 1Document66 pagesBlock 1HkgihijehNo ratings yet
- Cardio Quiz 1Document30 pagesCardio Quiz 1Dennis BaluyutNo ratings yet
- Chapter 11 - Endocrine SystemDocument13 pagesChapter 11 - Endocrine SystemIvy CustodioNo ratings yet
- Chapter 3 PerceptionDocument13 pagesChapter 3 PerceptionThavasi mari selvam NNo ratings yet
- The Molecular Cytoarchitecture of The Adult Mouse BrainDocument28 pagesThe Molecular Cytoarchitecture of The Adult Mouse BrainVictor Rodrigues SantosNo ratings yet
- Extra Credit Critical Thinking Questions: The Human Body, An OrientationDocument3 pagesExtra Credit Critical Thinking Questions: The Human Body, An OrientationFarzana KamilNo ratings yet
- Lojban ThesaurusDocument18 pagesLojban ThesaurusmarkrossoNo ratings yet
- LI Fakultas KedokteranDocument23 pagesLI Fakultas KedokteranChasnaaLidzaMalihaNo ratings yet
- Ethanol Tolerance in Bacteria PDFDocument2 pagesEthanol Tolerance in Bacteria PDFReneeNo ratings yet
- Femur: Anatomy of The KneeDocument2 pagesFemur: Anatomy of The KneeKetan MNo ratings yet
- Neet PG Mock 4Document65 pagesNeet PG Mock 4Diwakesh C B100% (1)
- Dialogue For Student NursesDocument2 pagesDialogue For Student NursesYopa YopisaNo ratings yet
- CH 06Document6 pagesCH 06Enjie Elrassi100% (1)
- GSRDocument10 pagesGSRNaman Kulshresth50% (2)
- Skin Lumps, Cyst, BumpsDocument6 pagesSkin Lumps, Cyst, BumpsMaria AgathaNo ratings yet
- "The Tao of Rejuvenation" Interview With Angelo DrudaDocument9 pages"The Tao of Rejuvenation" Interview With Angelo DrudagaryscottmorrowNo ratings yet
- Bio IA Possible TopicsDocument5 pagesBio IA Possible TopicsKirsten Yu100% (1)
- Flowering PlantDocument21 pagesFlowering PlantAhmed SharifNo ratings yet
- Anciden and Emergansi Exsample Ancident 1. FracturesDocument8 pagesAnciden and Emergansi Exsample Ancident 1. Fracturesaldys rosalindaNo ratings yet