Download as pdf or txt
You might also like
- Ultimate Grocery ListDocument1 pageUltimate Grocery ListGaston Wilson100% (13)
- Reproduction and Sexuality in Marine Fishes: Patterns and ProcessesFrom EverandReproduction and Sexuality in Marine Fishes: Patterns and ProcessesKathleen S. ColeNo ratings yet
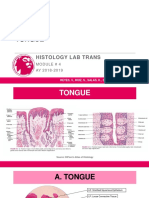
- 4.1A-Tongue (GIT)Document11 pages4.1A-Tongue (GIT)camille chua0% (1)
- Kindergarten Library Skills Unit PlanDocument6 pagesKindergarten Library Skills Unit PlanAmandaNo ratings yet
- Velez-Rubio Et Al. 2018 - Pre and Post-Settlement Movements of Juvenile Green Turtles in The Southwestern Atlantic OceanDocument10 pagesVelez-Rubio Et Al. 2018 - Pre and Post-Settlement Movements of Juvenile Green Turtles in The Southwestern Atlantic OceanKarumbéNo ratings yet
- Population Genetics and Phylogeography of Sea TurtlesDocument23 pagesPopulation Genetics and Phylogeography of Sea Turtleshunterdog42No ratings yet
- 1 PB PDFDocument10 pages1 PB PDFAlfa DarojatinNo ratings yet
- Markaida Et Al 2018 Reproduction - and - Population - Structure - of - Octopus - Mimus - EcuadorDocument7 pagesMarkaida Et Al 2018 Reproduction - and - Population - Structure - of - Octopus - Mimus - EcuadorAncistrocheirusNo ratings yet
- Symbiotic Association Between Caprellids (Amphipoda: Caprellidae) and The Scorpionfish Scorpaena Mystes (Pisces: Scorpaenidae)Document6 pagesSymbiotic Association Between Caprellids (Amphipoda: Caprellidae) and The Scorpionfish Scorpaena Mystes (Pisces: Scorpaenidae)CRISTIAN MOISES GALVAN VILLANo ratings yet
- Reproductive Behavior and Embryonic Development of The Pharaoh Cuttlefish, Sepia PharaonisDocument16 pagesReproductive Behavior and Embryonic Development of The Pharaoh Cuttlefish, Sepia Pharaonis李孟芳No ratings yet
- Landaeta Et Al 2011 Feeding Habits of Larval Maurolicus Parvipinnis in Patagonian FjordsDocument12 pagesLandaeta Et Al 2011 Feeding Habits of Larval Maurolicus Parvipinnis in Patagonian FjordsjoancoasNo ratings yet
- Jurnal CephalochordataDocument3 pagesJurnal CephalochordataAnna KareninaNo ratings yet
- Evolutionary Applications - 2019 - JenkinsDocument19 pagesEvolutionary Applications - 2019 - JenkinsIara GonçalvesNo ratings yet
- Biology: Characterization of Bottlenose Dolphin (Tursiops Truncatus) Sperm Based On Morphometric TraitsDocument17 pagesBiology: Characterization of Bottlenose Dolphin (Tursiops Truncatus) Sperm Based On Morphometric TraitsImas-hendryNo ratings yet
- Peces de La Laguna Cormorán, Parque Nacional Sangay, EcuadorDocument10 pagesPeces de La Laguna Cormorán, Parque Nacional Sangay, EcuadorTatiana GaonaNo ratings yet
- Dermochelys Coriacea SkeletonDocument13 pagesDermochelys Coriacea SkeletonYessica VargasNo ratings yet
- Artigo 1 PDFDocument10 pagesArtigo 1 PDFLara RodriguesNo ratings yet
- Sphyrna Lewini: Global Phylogeography of The Scalloped Hammerhead SharkDocument13 pagesSphyrna Lewini: Global Phylogeography of The Scalloped Hammerhead SharkRodrigo MazzoleniNo ratings yet
- Molluscan Studies: Journal ofDocument7 pagesMolluscan Studies: Journal ofJAVIER ANDRES VENEGAS OJEDANo ratings yet
- Romero-Caicedo2015 Raya GuitarraDocument15 pagesRomero-Caicedo2015 Raya GuitarraALHONDRA HORTENCIA GOMEZ SANCHEZNo ratings yet
- Residency and Abundance of Sperm Whales (Physeter Macrocephalus) in Nemuro Strait, Hokkaido, JapanDocument11 pagesResidency and Abundance of Sperm Whales (Physeter Macrocephalus) in Nemuro Strait, Hokkaido, JapanGianni PavanNo ratings yet
- Supérvivencia y LogebidadDocument10 pagesSupérvivencia y LogebidadDaniel VargasNo ratings yet
- 27 Segura EpocaDocument12 pages27 Segura EpocaVanessa Usbeck LópezNo ratings yet
- Phylogeography, Population History and Conservation Genetics of Jaguars (, Mammalia, Felidae)Document16 pagesPhylogeography, Population History and Conservation Genetics of Jaguars (, Mammalia, Felidae)Luis Alexandre - GaelNo ratings yet
- Artículo Redalyc 44943437032Document23 pagesArtículo Redalyc 44943437032Fernanda Charqueño CelisNo ratings yet
- Des Emb Pez AngelDocument7 pagesDes Emb Pez AngelJostin JoutexNo ratings yet
- Shillinger 2008 PlosDocument9 pagesShillinger 2008 PlostortugamarinaNo ratings yet
- Escherichia Coli Populations in Great Lakes Waterfowl Exhibit SpatialDocument6 pagesEscherichia Coli Populations in Great Lakes Waterfowl Exhibit SpatialMohamed Amin MazenNo ratings yet
- Preproposal FinalDocument9 pagesPreproposal Finalapi-254424729No ratings yet
- Biology and Blooms of The Edible Jellyfish in The Saleh Bay, IndonesiaDocument14 pagesBiology and Blooms of The Edible Jellyfish in The Saleh Bay, Indonesiabuya301290No ratings yet
- TMP 2 CCDocument13 pagesTMP 2 CCFrontiersNo ratings yet
- Artículo de RenacuajosDocument9 pagesArtículo de RenacuajosAndres Felipe BermudezNo ratings yet
- A1 PDFDocument11 pagesA1 PDFwulanNo ratings yet
- 0034 7744 RBT 63 s2 183Document12 pages0034 7744 RBT 63 s2 183Katherine A AlcNo ratings yet
- TMP DBA0Document15 pagesTMP DBA0FrontiersNo ratings yet
- FSR 175Document10 pagesFSR 175Michaiel PiticarNo ratings yet
- Guidetti 2013Document4 pagesGuidetti 2013Fadhli LatuconsinaNo ratings yet
- Besouros de Água Mo Et Al.Document20 pagesBesouros de Água Mo Et Al.thaisa christineNo ratings yet
- Diversity and Dispersion Patterns of Echinoderms in Babanlagan, Talisayan, Misamis Oriental, PhilippinesDocument5 pagesDiversity and Dispersion Patterns of Echinoderms in Babanlagan, Talisayan, Misamis Oriental, PhilippinesGeloWalagNo ratings yet
- Spatial Distribution of Cephalopod Paralarvae in San José Gulf (Northern Patagonia, Argentina) : The Role of Tidal Circulation in Larval DispersalDocument8 pagesSpatial Distribution of Cephalopod Paralarvae in San José Gulf (Northern Patagonia, Argentina) : The Role of Tidal Circulation in Larval DispersallujanvgNo ratings yet
- Estructura Trófica de Los Peces en Arroyos Del Corral de San Luis, Cuenca Del Bajo Magdalena, Caribe, ColombiaDocument19 pagesEstructura Trófica de Los Peces en Arroyos Del Corral de San Luis, Cuenca Del Bajo Magdalena, Caribe, ColombiaBaltazar Calderon RoncalloNo ratings yet
- Mexican Dancer in Ecuador: Molecular Confirmation, Embryology and Planktotrophy in The Sea Slug Elysia DiomedeaDocument30 pagesMexican Dancer in Ecuador: Molecular Confirmation, Embryology and Planktotrophy in The Sea Slug Elysia DiomedeaLola MoreanoNo ratings yet
- The Reproductive Ecology of Exotic Trachemys Scripta Elegans in An Invaded Area of Southern EuropeDocument10 pagesThe Reproductive Ecology of Exotic Trachemys Scripta Elegans in An Invaded Area of Southern Europewijayanti chantikaNo ratings yet
- Species Diversity and Spatial Structure of Intertidal Mollusks in Padada, Davao Del Sur, PhilippinesDocument10 pagesSpecies Diversity and Spatial Structure of Intertidal Mollusks in Padada, Davao Del Sur, PhilippinesYayan MardiansyahNo ratings yet
- 813 1308 1 PBDocument11 pages813 1308 1 PBDombou RolandNo ratings yet
- Freshwater EcologyDocument12 pagesFreshwater EcologyMaricris I. AbuanNo ratings yet
- Overview of The Order Zoantharia (Cnidaria: Anthozoa) in BrazilDocument13 pagesOverview of The Order Zoantharia (Cnidaria: Anthozoa) in BrazilRabha EnanehcNo ratings yet
- Mitogenomes Uncover Extinct Penguin Taxa and Reveal Island Formation As A Key Driver of SpeciationDocument14 pagesMitogenomes Uncover Extinct Penguin Taxa and Reveal Island Formation As A Key Driver of SpeciationayunikmatulNo ratings yet
- Early Ontogenesis of The Angelfish, Pterophyllum Scalare Schultze, 1823 (Cichlidae)Document10 pagesEarly Ontogenesis of The Angelfish, Pterophyllum Scalare Schultze, 1823 (Cichlidae)kusuma wardanaNo ratings yet
- 52 89 1 PBDocument15 pages52 89 1 PBsister girlfriendNo ratings yet
- Word Materi 9Document13 pagesWord Materi 9santiNo ratings yet
- Galimany Et Al. 2015Document10 pagesGalimany Et Al. 2015Marco Antonio SolisNo ratings yet
- 2012 Ecological - ISMEDocument14 pages2012 Ecological - ISMEVERONICA BLANCONo ratings yet
- Eugerres MexicanusDocument12 pagesEugerres MexicanusFlavioNo ratings yet
- Pliego-Cardenas Et Al. 2020 Genetic Structure of Octopus MimusDocument9 pagesPliego-Cardenas Et Al. 2020 Genetic Structure of Octopus MimusAncistrocheirusNo ratings yet
- Estructura Trófica de Los Peces en Arroyos Del Corral de San Luis, Cuenca Del Bajo Magdalena, Caribe, ColombiaDocument18 pagesEstructura Trófica de Los Peces en Arroyos Del Corral de San Luis, Cuenca Del Bajo Magdalena, Caribe, ColombiaKate HowardNo ratings yet
- Rodrigues Galdino2009Document14 pagesRodrigues Galdino2009Belay AbdissaNo ratings yet
- Habitos Alimentarios de Prionace GlaucaDocument13 pagesHabitos Alimentarios de Prionace GlaucaJorge BarturenNo ratings yet
- Feedinghabits 2Document9 pagesFeedinghabits 2egha trishnayanaNo ratings yet
- Estructuratrficadelospecesenarroyosdel Corralde San Luis 2016Document21 pagesEstructuratrficadelospecesenarroyosdel Corralde San Luis 2016Baltazar Calderon RoncalloNo ratings yet
- Orcinus Orca EcuadorDocument11 pagesOrcinus Orca EcuadorMiguel90No ratings yet
- Early Development of Betta Splendens UndDocument10 pagesEarly Development of Betta Splendens UndIsabella ValenciaNo ratings yet
- PDF 26Document12 pagesPDF 26IVAN ANDRESNo ratings yet
- Treasure Hunters Book I Players' GuideDocument44 pagesTreasure Hunters Book I Players' Guidewoem4985100% (1)
- Menstrual Cycle Associated Changes in Agility in Amateur Female AthletesDocument39 pagesMenstrual Cycle Associated Changes in Agility in Amateur Female Athletesvishnu priyaNo ratings yet
- Bengal Florican (Houbaropsis Bengalensis)Document19 pagesBengal Florican (Houbaropsis Bengalensis)effaNo ratings yet
- ExercisesDocument11 pagesExercisesBechara HaddadNo ratings yet
- Dogs British English Student Ver2Document3 pagesDogs British English Student Ver2Josef ŠtaifNo ratings yet
- Classic D&D Random Encounter Table & Tres Gen Compiled EditionDocument120 pagesClassic D&D Random Encounter Table & Tres Gen Compiled EditionRonHowardNo ratings yet
- Billy Boy-Terry GreenwellDocument21 pagesBilly Boy-Terry GreenwellМАРИЯ ПетковаNo ratings yet
- General Tamil Part - A, B & CDocument75 pagesGeneral Tamil Part - A, B & Cabi99lashNo ratings yet
- 990916 EUS教學 (14) 急診超音波在胸腔急症之應用Document63 pages990916 EUS教學 (14) 急診超音波在胸腔急症之應用juice119No ratings yet
- The Food IndustryDocument28 pagesThe Food IndustryIleana Violeta CarmocanuNo ratings yet
- Critical Thinking On Certain Questions Concerning Comparative Anatomy of The VertebratesDocument5 pagesCritical Thinking On Certain Questions Concerning Comparative Anatomy of The VertebratesSeth Andrew SalihNo ratings yet
- Resume 1Document1 pageResume 1api-336472593No ratings yet
- Anatomy and Physiology of Child Respiratory SystemDocument8 pagesAnatomy and Physiology of Child Respiratory Systemmanju talluriNo ratings yet
- Oral Histology Odontoword Puzzle Clues & AnswersDocument2 pagesOral Histology Odontoword Puzzle Clues & Answersdr arjunNo ratings yet
- Proto-Nostratic VocabularyDocument82 pagesProto-Nostratic VocabularyProfessor Valdemir Siqueira da SilvaNo ratings yet
- 1 # Accounting For The DeadDocument103 pages1 # Accounting For The DeadMateusz BiałekNo ratings yet
- Reproductive Biology McqsDocument7 pagesReproductive Biology McqsAyan Mukherjee100% (1)
- ADocument80 pagesAhowardalistairroarkNo ratings yet
- An Atlas of Radiology of The Traumatized Dog and CatDocument566 pagesAn Atlas of Radiology of The Traumatized Dog and CatDenise Gomes de MeloNo ratings yet
- Reflexes That Control Cardiovascular FunctionDocument11 pagesReflexes That Control Cardiovascular FunctionRaam NaikNo ratings yet
- Cockfighting State LawsDocument4 pagesCockfighting State LawsOur CompassNo ratings yet
- Dr. Jeff - Rocky Mountain Vet S2 Ep 01Document82 pagesDr. Jeff - Rocky Mountain Vet S2 Ep 01Varga KrisztaNo ratings yet
- Wod LichDocument6 pagesWod LichDarkrahodNo ratings yet
- The Happy Mirror StoryDocument8 pagesThe Happy Mirror StoryEmmanuel RecodoNo ratings yet
- Nomenclatura ParásitosDocument28 pagesNomenclatura ParásitosAylinNayeliNo ratings yet
- QuestDocument16 pagesQuestAbhishek WalNo ratings yet
- Hygiene and Sanitation Ms-1Document26 pagesHygiene and Sanitation Ms-1genesisrevelations100% (1)