Download as pdf or txt
You might also like
- BHG Beginner Garden For Shade Garden PlanDocument2 pagesBHG Beginner Garden For Shade Garden PlanDaniela BracchittaNo ratings yet
- Generic Organic Aquaponics Business Plan 102111Document17 pagesGeneric Organic Aquaponics Business Plan 102111illanizaharudinNo ratings yet
- Germinacion de Semillas de Cuatro Especies ArboreaDocument15 pagesGerminacion de Semillas de Cuatro Especies Arboreajeniffer diazNo ratings yet
- Fenologia Das Árvores e Disponibilidade de Água em Sistemas Agroflorestais SemiáridosDocument13 pagesFenologia Das Árvores e Disponibilidade de Água em Sistemas Agroflorestais SemiáridosMarcelo da Silva IrmãoNo ratings yet
- 10.1023@A@1006063010677 Trema MicranthaDocument8 pages10.1023@A@1006063010677 Trema MicranthaClaudio SantanaNo ratings yet
- Miranda 2005Document11 pagesMiranda 2005Ariadne Cristina De AntonioNo ratings yet
- Comparative Water Relations of Mature Mahogany (Swietenia Dry Forest in BoliviaDocument8 pagesComparative Water Relations of Mature Mahogany (Swietenia Dry Forest in BoliviaVodoo RigbyNo ratings yet
- Review of Related LiteratureDocument12 pagesReview of Related LiteratureEllaNatividad0% (1)
- Ceccon Et Al Potencial Biológico 4 Esp Nativas Bosque Seco para RestauraciónDocument13 pagesCeccon Et Al Potencial Biológico 4 Esp Nativas Bosque Seco para RestauraciónAlejandro GilNo ratings yet
- Field Study of Beech (Fagus Sylvatica L.) and Melojo Oak (Quercus Pyrenaica Willd) Leaf Litter Decomposition in The Centre of The Iberian PeninsulaDocument12 pagesField Study of Beech (Fagus Sylvatica L.) and Melojo Oak (Quercus Pyrenaica Willd) Leaf Litter Decomposition in The Centre of The Iberian PeninsulaDiogo AraujoNo ratings yet
- Phytoliths As Indicators of Pedogenesis and Paleoenvironmental Changes in The Brazilian CerradoDocument5 pagesPhytoliths As Indicators of Pedogenesis and Paleoenvironmental Changes in The Brazilian Cerradokaname10No ratings yet
- Effects of Fire On Vegetation and Small Mammal Communities in A Mojave DesertDocument10 pagesEffects of Fire On Vegetation and Small Mammal Communities in A Mojave DesertRubens HayekNo ratings yet
- Guariguata,2000Document10 pagesGuariguata,2000alis23.artNo ratings yet
- Seed Germination Conditions and Implications For Establishment of An Epiphyte, Aechmea Bracteata (Bromeliaceae)Document10 pagesSeed Germination Conditions and Implications For Establishment of An Epiphyte, Aechmea Bracteata (Bromeliaceae)José David QuevedoNo ratings yet
- Flora and Life-Form Spectrum in An Area of Deciduous Thorn Woodland (Caatinga) in Northeastern, BrazilDocument11 pagesFlora and Life-Form Spectrum in An Area of Deciduous Thorn Woodland (Caatinga) in Northeastern, BrazilCecili MendesNo ratings yet
- Trees23 787Document13 pagesTrees23 787Sandra Arroyo AlfaroNo ratings yet
- Austral Ecology - 2008 - Lusk - Survival and Growth of Seedlings of 12 Chilean Rainforest Trees in Two Light EnvironmentsDocument10 pagesAustral Ecology - 2008 - Lusk - Survival and Growth of Seedlings of 12 Chilean Rainforest Trees in Two Light EnvironmentsAlén MirandaNo ratings yet
- Chapter IiDocument7 pagesChapter IicreshafayetteNo ratings yet
- Seedling Grow VALIODocument9 pagesSeedling Grow VALIOsemirian_amoedo3994No ratings yet
- Martinez - Garza 2011 Dispersión de Semillas PDFDocument12 pagesMartinez - Garza 2011 Dispersión de Semillas PDFVanessa AraujoNo ratings yet
- Becerra Et Al. 2011 Efecto HierbasDocument9 pagesBecerra Et Al. 2011 Efecto HierbasAntonia Gómez BaginskyNo ratings yet
- Pennisetum PurpureumDocument3 pagesPennisetum PurpureumHildan H GNo ratings yet
- Seedling Functional Types in A Lowland Rain Forest in MexicoDocument13 pagesSeedling Functional Types in A Lowland Rain Forest in MexicoJim VillenaNo ratings yet
- Lopez Iglesias 2014Document9 pagesLopez Iglesias 2014Norberto JúniorNo ratings yet
- Asquith e Chang 2005 - DSDocument12 pagesAsquith e Chang 2005 - DSBeatrizNo ratings yet
- Forest Ecology and Management: Post-Fire Tree Mortality in Mixed Forests of Central PortugalDocument9 pagesForest Ecology and Management: Post-Fire Tree Mortality in Mixed Forests of Central PortugalDitaChiquelaNo ratings yet
- Esclerofilia in Qualea Parviflora PDFDocument10 pagesEsclerofilia in Qualea Parviflora PDFHenrique VianaNo ratings yet
- Camargo Et Al 2018 BiotropicaDocument13 pagesCamargo Et Al 2018 BiotropicaAriadne Cristina De AntonioNo ratings yet
- 2003 LovelockDocument12 pages2003 LovelockMaristela VolpatoNo ratings yet
- MacroninvertebradosDocument20 pagesMacroninvertebradosdanielvargas1No ratings yet
- Lifetime Growth Patterns and Ages of Bolivian Rainforest Trees Obtained by Tree Ring AnalysisDocument18 pagesLifetime Growth Patterns and Ages of Bolivian Rainforest Trees Obtained by Tree Ring AnalysisCatalin RoibuNo ratings yet
- 10 1016@j Scitotenv 2017 04 100 PDFDocument16 pages10 1016@j Scitotenv 2017 04 100 PDFfiqriNo ratings yet
- Allelopathy - A Tool For Weed Management in ForestDocument15 pagesAllelopathy - A Tool For Weed Management in ForestsaporettiNo ratings yet
- 5780 8073 1 PBDocument18 pages5780 8073 1 PBJesús ArroyoNo ratings yet
- GiraldoValle CativoDocument20 pagesGiraldoValle CativoClaudia ParadaNo ratings yet
- Monteiro Et Al 2013 Dissolved Organic Carbon ConcentrationDocument7 pagesMonteiro Et Al 2013 Dissolved Organic Carbon ConcentrationInaura SantosNo ratings yet
- Conocarpus ErectusDocument3 pagesConocarpus ErectusLa ChispaNo ratings yet
- Nitrogen-Fixing Legume Tree Species For The Reclamation of Severely Degraded Lands in BrazilDocument11 pagesNitrogen-Fixing Legume Tree Species For The Reclamation of Severely Degraded Lands in BrazilFadhlan Husnul HafizhNo ratings yet
- 2248 7026 Rfnam 72 01 08751Document12 pages2248 7026 Rfnam 72 01 08751Andrés BoteroNo ratings yet
- A 26 V 60 N 2Document14 pagesA 26 V 60 N 2Diego Andres Moreno GaonaNo ratings yet
- DCA - 5 - Marginal HollyDocument9 pagesDCA - 5 - Marginal HollyGuillermo BuragliaNo ratings yet
- Diversity of Vascular Plant Species in An Agroforest: The Case of A Rubber (Hevea Brasiliensis) Plantation in Makilala, North CotabatoDocument8 pagesDiversity of Vascular Plant Species in An Agroforest: The Case of A Rubber (Hevea Brasiliensis) Plantation in Makilala, North CotabatoMary Joy ManayagaNo ratings yet
- NascimentoDocument11 pagesNascimentotaucci123No ratings yet
- Tree Species Diversity, Composition and Aboveground Biomass Across Dry Forest Land-Cover Types in Coastal EcuadorDocument13 pagesTree Species Diversity, Composition and Aboveground Biomass Across Dry Forest Land-Cover Types in Coastal EcuadorLouis Paul KABELONGNo ratings yet
- Guevara 2005Document8 pagesGuevara 2005Roger GuevaraNo ratings yet
- Patogeni Quercia Da Sughero - Ragazzi Et AlDocument10 pagesPatogeni Quercia Da Sughero - Ragazzi Et AlmncmarcoNo ratings yet
- Effect of Drought Stress On Growth and Flowering of Marigold (Tagetes Erecta L.)Document9 pagesEffect of Drought Stress On Growth and Flowering of Marigold (Tagetes Erecta L.)Dheeraj VermaNo ratings yet
- Cacti: Notes On Their Uses and Potential For Climate Change MitigationDocument17 pagesCacti: Notes On Their Uses and Potential For Climate Change MitigationJohao Girón AzañeroNo ratings yet
- Diversity and Floristic Composition of Neotropical Dry ForestsDocument43 pagesDiversity and Floristic Composition of Neotropical Dry ForestsJosé HernándezNo ratings yet
- Exotic Plant Species Invade Hot Spots of Native Plant DiversityDocument22 pagesExotic Plant Species Invade Hot Spots of Native Plant DiversitysellaginellaNo ratings yet
- Identi®cation of Conservation Measures To Protect The Japanese Endangered Plant Species Aster KantoensisDocument9 pagesIdenti®cation of Conservation Measures To Protect The Japanese Endangered Plant Species Aster Kantoensismaria bNo ratings yet
- Mendes Et Al. 2017 - Reproductive Phenology of Mauritia Flexuosa L. (Arecaceae)Document9 pagesMendes Et Al. 2017 - Reproductive Phenology of Mauritia Flexuosa L. (Arecaceae)Yakov Mario QuinterosNo ratings yet
- Alfalfa Anegamiento FenotiposDocument9 pagesAlfalfa Anegamiento Fenotiposcamilajaime1993No ratings yet
- Papel de Los Mamíferos en Los Procesos de Dispersión y Depredación de Semillas de Mauritia Flexuosa (Arecaceae) en La..Document13 pagesPapel de Los Mamíferos en Los Procesos de Dispersión y Depredación de Semillas de Mauritia Flexuosa (Arecaceae) en La..Mariana Gutierrez MuneraNo ratings yet
- Mares ErnstDocument20 pagesMares ErnstJoana FerreiraNo ratings yet
- Pinus Radiata D. Don Subject To Drought: Impact of Mycorrhizae and Irrigation in The Survival of Seedlings ofDocument9 pagesPinus Radiata D. Don Subject To Drought: Impact of Mycorrhizae and Irrigation in The Survival of Seedlings ofFranco GianniniNo ratings yet
- Diversity of Soil Fauna and Ecosystem FunctionDocument14 pagesDiversity of Soil Fauna and Ecosystem Functiontariqulmasud12No ratings yet
- Article Stomatal Density Leaf Area of RhizophoraDocument13 pagesArticle Stomatal Density Leaf Area of Rhizophorasara.najjariNo ratings yet
- Lewis - 2000 - Selective Logging ButterfliesDocument12 pagesLewis - 2000 - Selective Logging ButterfliesKellyta RodriguezNo ratings yet
- Seedling Growth of Fifteen Brazilian Tropical TreeDocument13 pagesSeedling Growth of Fifteen Brazilian Tropical TreeMarina Pérola ZerbinatoNo ratings yet
- Ministry of Education Fiji Year 13 Certificate Examination 2017Document45 pagesMinistry of Education Fiji Year 13 Certificate Examination 2017C3real KillerNo ratings yet
- Ibn Sina.Document4 pagesIbn Sina.Ubaidillah YusofNo ratings yet
- Intergrated Science Grade 9Document16 pagesIntergrated Science Grade 9Prof Samuel Kashina100% (1)
- A. Delonix Regia: Answer All Questions in This SectionDocument9 pagesA. Delonix Regia: Answer All Questions in This SectionNorbert LimNo ratings yet
- A Detailed Lesson Plan in Grade 10Document215 pagesA Detailed Lesson Plan in Grade 10Chermaine VillanuevaNo ratings yet
- Activity Sheet: Activity Sheet Number 2 Classification and Identification of CropsDocument8 pagesActivity Sheet: Activity Sheet Number 2 Classification and Identification of CropsLocelTanJavillonarNo ratings yet
- Humidity and Light Affect The Growth, Development and Nitrogenase Activity of Stem Nodules of Sesbania Rostrata BremDocument8 pagesHumidity and Light Affect The Growth, Development and Nitrogenase Activity of Stem Nodules of Sesbania Rostrata BremNdra PompomorinNo ratings yet
- Dps GNGR Class 1 Evs NotesDocument72 pagesDps GNGR Class 1 Evs NotesPRNo ratings yet
- Evolving Concept of Life Based On Emerging PiecesDocument25 pagesEvolving Concept of Life Based On Emerging PiecesPierre Suayan100% (1)
- Plant Competition in A Changing WorldDocument156 pagesPlant Competition in A Changing WorldMona RamosNo ratings yet
- Test Ket 8Document4 pagesTest Ket 8momobear152No ratings yet
- Interior Plants: Selection and Care: Lecturer, Plant SciencesDocument7 pagesInterior Plants: Selection and Care: Lecturer, Plant SciencessabumathewNo ratings yet
- ANNEXURE I List of Indian InstitutionsDocument82 pagesANNEXURE I List of Indian InstitutionsFrancis MwangiNo ratings yet
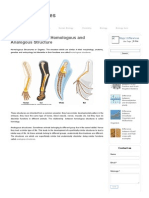
- Difference Between Homologous and Analogous Structure - Major DifferencesDocument3 pagesDifference Between Homologous and Analogous Structure - Major DifferencesArvind BoudhaNo ratings yet
- Grade 11 Life Sciences Remote Learning Booklet - Term 1Document56 pagesGrade 11 Life Sciences Remote Learning Booklet - Term 1lungstachapterNo ratings yet
- Horticultural Classification Terms: CMG Gardennotes #121Document8 pagesHorticultural Classification Terms: CMG Gardennotes #121Mohd Khairil Hilmi HalimNo ratings yet
- B2 Questions and AnswersDocument22 pagesB2 Questions and Answersinaayairshad.17No ratings yet
- Bio EngineeringDocument1 pageBio EngineeringShuvanjan Dahal100% (1)
- Science Planning-HabitatsDocument8 pagesScience Planning-Habitatsapi-300499643No ratings yet
- Clss 11Document26 pagesClss 11Nisarg PatelNo ratings yet
- Hydroponic Nutrient Solution - The Essential GuideDocument10 pagesHydroponic Nutrient Solution - The Essential GuideRómulo Del ValleNo ratings yet
- S Stage 9 p110 Ms Afp PDFDocument16 pagesS Stage 9 p110 Ms Afp PDFpixelhoboNo ratings yet
- Strawberry JournalDocument12 pagesStrawberry JournalJoydeb GomastaNo ratings yet
- Byjus Com Biology Plant PhysiologyDocument14 pagesByjus Com Biology Plant PhysiologyVikas PratikNo ratings yet
- Bio Grade 9 Revision Semester 1Document16 pagesBio Grade 9 Revision Semester 1GeoHimselfNo ratings yet
- 2,4-D and Tomato YieldDocument5 pages2,4-D and Tomato YieldtasuafNo ratings yet
- A Beginning Farmer 'S Guide To Agriculture Words, Terms and DefinitionsDocument12 pagesA Beginning Farmer 'S Guide To Agriculture Words, Terms and DefinitionsaaNo ratings yet
- Bio 2.2 EcosystemDocument7 pagesBio 2.2 EcosystemSuzuko YukinoNo ratings yet