Download as pdf or txt
You might also like
- Biohemija Skripta FocaDocument43 pagesBiohemija Skripta FocaNevena JovićNo ratings yet
- Obrada Primarnog TranskriptaDocument13 pagesObrada Primarnog TranskriptaGoranNo ratings yet
- Pitanja Iz Celijske Biologije 1Document11 pagesPitanja Iz Celijske Biologije 1davidmarkovic032No ratings yet
- БИОСИНТЕЗА ПРОТЕИНА 1Document15 pagesБИОСИНТЕЗА ПРОТЕИНА 1Musika ArtoNo ratings yet
- TRANSKRIPCIJADocument8 pagesTRANSKRIPCIJAmilena zugićNo ratings yet
- Gen Genom I ReplikacijaDocument53 pagesGen Genom I ReplikacijaDejan Mitic100% (2)
- GIM 3. Ekspresija Gena 2017Document60 pagesGIM 3. Ekspresija Gena 2017Nevena TodosijevicNo ratings yet
- Biljni Hormoni 3Document39 pagesBiljni Hormoni 3GoranNo ratings yet
- Значај воде за живот биљакаDocument39 pagesЗначај воде за живот биљакаGoran100% (1)
- Homeostaza, Osetljivost, Pokretljivost, PMDocument16 pagesHomeostaza, Osetljivost, Pokretljivost, PMGoranNo ratings yet
- VarenjeDocument48 pagesVarenjeGoranNo ratings yet
- TranskripcijaDocument21 pagesTranskripcijaGoranNo ratings yet
- Predavanje - Vezani Geni I RekombinacijeDocument41 pagesPredavanje - Vezani Geni I RekombinacijeDoraNo ratings yet
- Nasledjivanje Pola. Promene U Strukturi I Broju HromozomaDocument22 pagesNasledjivanje Pola. Promene U Strukturi I Broju HromozomaGoranNo ratings yet
- Biosinteza Proteina TranslacijaDocument22 pagesBiosinteza Proteina TranslacijaGoran100% (1)
- 06 Molekularna BiotehnolDocument25 pages06 Molekularna BiotehnolKlaudia KamraNo ratings yet
- Metabolizam Pojam I Principi 2Document9 pagesMetabolizam Pojam I Principi 2GoranNo ratings yet
- ENZIMIDocument17 pagesENZIMIGoranNo ratings yet
- Krebsov CiklusDocument16 pagesKrebsov CiklusGoranNo ratings yet
- 9Document28 pages9Vladimir VelimirovicNo ratings yet
- Metode Molekularne GenetikeDocument16 pagesMetode Molekularne GenetikeCíntia 20 MuenhoNo ratings yet
- 4 Antitela I AntigeniDocument56 pages4 Antitela I AntigeniIlakiNo ratings yet
- Проток Информација у Живом Свету - ДНК и РНКDocument34 pagesПроток Информација у Живом Свету - ДНК и РНКGoran100% (1)
- Disanje GlikolizaDocument12 pagesDisanje GlikolizaGoranNo ratings yet
- JedroDocument15 pagesJedroSofija ŽivkovićNo ratings yet
- Adaptacije biljaкa na dejstvo eкološкih faкtoraDocument29 pagesAdaptacije biljaкa na dejstvo eкološкih faкtoraGoranNo ratings yet
- OplodjenjeDocument13 pagesOplodjenjeGoranNo ratings yet
- ТРАНСКРИПЦИЈА, ОБРАДА ПРИМАРНОГ ТРАНСКРИПТАDocument33 pagesТРАНСКРИПЦИЈА, ОБРАДА ПРИМАРНОГ ТРАНСКРИПТАGoran0% (1)
- Građa Ćelijske Membrane 2019 ScribdDocument20 pagesGrađa Ćelijske Membrane 2019 ScribdGoranNo ratings yet
- Celijska Komunikacija 2022 ScribdDocument28 pagesCelijska Komunikacija 2022 ScribdGoranNo ratings yet
- Transkripcijamatrična Sinteza iRNK 169Document21 pagesTranskripcijamatrična Sinteza iRNK 169Goran100% (1)
- Uredjaj Za Proveravanje Bernulijeve JednacineDocument4 pagesUredjaj Za Proveravanje Bernulijeve JednacineАлександар ДимитријевићNo ratings yet
- VodaDocument14 pagesVodaGoranNo ratings yet
- Molekularna Biologija 1 2015Document83 pagesMolekularna Biologija 1 2015GoranNo ratings yet
- Regulacija Aktivnosti Gena Kod ProkariotaDocument30 pagesRegulacija Aktivnosti Gena Kod Prokariota4ndrejNo ratings yet
- РЕПЛИКАЦИЈАDocument4 pagesРЕПЛИКАЦИЈАAndrea KovačevićNo ratings yet
- Organizacija Humanog GenomaDocument7 pagesOrganizacija Humanog GenomaHexaGlobinNo ratings yet
- Celijske OrganeleDocument2 pagesCelijske OrganeleLela SNo ratings yet
- Homeostaza, Osetljivost, Pokretljivost DJDocument11 pagesHomeostaza, Osetljivost, Pokretljivost DJGoran100% (1)
- Osnovna Pravila NasleđivanjaDocument23 pagesOsnovna Pravila NasleđivanjaGoranNo ratings yet
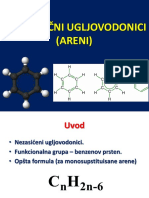
- Aromati, PH (1) 8133096057255525017 PDFDocument80 pagesAromati, PH (1) 8133096057255525017 PDFDamnjanNo ratings yet
- Odgovori Na Pitanja Genetika Kolokvijum LDocument12 pagesOdgovori Na Pitanja Genetika Kolokvijum LMarija KarkićNo ratings yet
- Enzimi PMDocument20 pagesEnzimi PMGoran100% (1)
- Čulni Organi DR Krstic VladimirDocument58 pagesČulni Organi DR Krstic Vladimirdejan0705No ratings yet
- Glavna SkriptaDocument83 pagesGlavna SkriptaMohanad YoByNo ratings yet
- Culni SistemDocument20 pagesCulni SistemDejan PuhaloNo ratings yet
- Koenzimi 4Document71 pagesKoenzimi 4Rastko ZečevićNo ratings yet
- Genetički KodDocument8 pagesGenetički KodGoran100% (2)
- Evolucija I Organizacija Genoma Čoveka MajaDocument17 pagesEvolucija I Organizacija Genoma Čoveka MajaMikica HaalilovicNo ratings yet
- Osnovi Imunologije MDocument7 pagesOsnovi Imunologije MNikola PiljicNo ratings yet
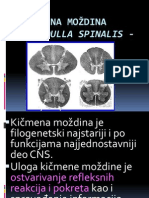
- Kicmena MozdinaDocument16 pagesKicmena MozdinaMicoouNo ratings yet
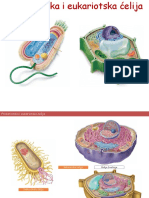
- Prokariotska I Eukariotska ĆelijaDocument10 pagesProkariotska I Eukariotska ĆelijaGoranNo ratings yet
- Biologija ĆelijeDocument6 pagesBiologija ĆelijeMilford11No ratings yet
- Oksidativna FosforilacijaDocument64 pagesOksidativna FosforilacijaИва ИлићNo ratings yet
- T Limfociti: Markeri I RazvojDocument34 pagesT Limfociti: Markeri I RazvojVidak ČovićNo ratings yet
- Ćelijska Gradja, Rast I RazvićeDocument12 pagesĆelijska Gradja, Rast I RazvićeGoranNo ratings yet
- 6 BCR I TCR Kompleks - PPT Sazrevanje T I B LimfocitaDocument30 pages6 BCR I TCR Kompleks - PPT Sazrevanje T I B LimfocitaBehireta Cimirotić DautbegovićNo ratings yet
- Molekularni Mehanizmi Lučenja Sekreta GIT I Regulacije ApetitaDocument55 pagesMolekularni Mehanizmi Lučenja Sekreta GIT I Regulacije ApetitaPavle BoskovicNo ratings yet
- Skripta Iz BiologijeDocument178 pagesSkripta Iz BiologijeDora MeštrovićNo ratings yet
- 17.biosinteza Proteina - 2019Document14 pages17.biosinteza Proteina - 2019Igor Mallix BukvaNo ratings yet
- Najcesce Nepravilnosti U Popunjavanju Potvrda o SmrtiDocument26 pagesNajcesce Nepravilnosti U Popunjavanju Potvrda o SmrtileagagaNo ratings yet
- Doktorske TituleDocument2 pagesDoktorske TituleleagagaNo ratings yet
- Nuklearna Magnetna RezonancaDocument16 pagesNuklearna Magnetna RezonancaleagagaNo ratings yet
- Najcesce Nepravilnosti U Popunjavanju Potvrda o SmrtiDocument26 pagesNajcesce Nepravilnosti U Popunjavanju Potvrda o SmrtileagagaNo ratings yet
- Izvestaj EMC IspitivanjaDocument10 pagesIzvestaj EMC IspitivanjaleagagaNo ratings yet
- Kardioresinhronizacionna Terapija - CRT PDFDocument5 pagesKardioresinhronizacionna Terapija - CRT PDFleagagaNo ratings yet
- Uslovi Kojima Poverilac Stiče Pravo Raskida UgovoraDocument14 pagesUslovi Kojima Poverilac Stiče Pravo Raskida UgovoraleagagaNo ratings yet
- DMSA, Bubrezi SnimanjeDocument56 pagesDMSA, Bubrezi SnimanjeleagagaNo ratings yet
- NMR - RadDocument62 pagesNMR - RadleagagaNo ratings yet
- Jonizacija LekovaDocument25 pagesJonizacija LekovaleagagaNo ratings yet
- Poliklonska I Monoklonska AntitelaDocument46 pagesPoliklonska I Monoklonska AntitelaleagagaNo ratings yet
- Izrada Preparata Za MikroskopiranjeDocument18 pagesIzrada Preparata Za MikroskopiranjeleagagaNo ratings yet
- Neuronske MrezeDocument20 pagesNeuronske MrezeleagagaNo ratings yet
- Zahtevi Standarda Za AkreditacijuDocument66 pagesZahtevi Standarda Za AkreditacijuleagagaNo ratings yet
- Procena Kvaliteta Zivota (QOL)Document105 pagesProcena Kvaliteta Zivota (QOL)leagagaNo ratings yet
- Sistem KomplementaDocument30 pagesSistem KomplementaleagagaNo ratings yet
- Celijska MembranaDocument5 pagesCelijska MembranaAnaNikolovNo ratings yet
- Genetičke Razlike Između Čoveka I ŠimpanzeDocument20 pagesGenetičke Razlike Između Čoveka I ŠimpanzeleagagaNo ratings yet
- Procedura RadaDocument29 pagesProcedura Radaleagaga100% (1)