Download as pdf or txt
You might also like
- Arcoplate ALLSTEINT FayreDocument4 pagesArcoplate ALLSTEINT Fayrevampiry1972No ratings yet
- E 107Document15 pagesE 107Thomas MelgarNo ratings yet
- 30 Traits of An EmpathDocument5 pages30 Traits of An EmpathFangedNimphNo ratings yet
- Malaria Pathogenesis: Louis H. Miller, Michael F. Good, Genevieve MilonDocument6 pagesMalaria Pathogenesis: Louis H. Miller, Michael F. Good, Genevieve MilonElroy Magnus QuinlanNo ratings yet
- Fimmu 11 571816Document20 pagesFimmu 11 571816Ricardo GomezNo ratings yet
- Zii2043 PDFDocument10 pagesZii2043 PDFPablo VelazquezNo ratings yet
- Fcimb 03 00106Document9 pagesFcimb 03 00106api-272542020No ratings yet
- Malaria Immunology 71Document7 pagesMalaria Immunology 71Firda DamiruNo ratings yet
- Salmonella Typhi: From A Human Pathogen To A Vaccine Vector: Cellular & Molecular Immunology May 2008Document8 pagesSalmonella Typhi: From A Human Pathogen To A Vaccine Vector: Cellular & Molecular Immunology May 2008noordin MukasaNo ratings yet
- Pathogenesis of Leptospirosis Important Issues 2161 0703.1000e127 1Document2 pagesPathogenesis of Leptospirosis Important Issues 2161 0703.1000e127 1Anonymous 7jKR9XbNo ratings yet
- PIIS009286741631008XDocument15 pagesPIIS009286741631008XJatin GoyalNo ratings yet
- Chourasia Et Al 2020 Plasmodium Falciparum Clag9 Associated Pfrhoph Complex Is Involved in Merozoite Binding To HumanDocument14 pagesChourasia Et Al 2020 Plasmodium Falciparum Clag9 Associated Pfrhoph Complex Is Involved in Merozoite Binding To HumanArunaditya Deshmukh (Adi)No ratings yet
- Humanized Mouse Models For The Study of Human Malaria Parasite Biology, Pathogenesis, and ImmunityDocument10 pagesHumanized Mouse Models For The Study of Human Malaria Parasite Biology, Pathogenesis, and ImmunityAnisah MahardianiNo ratings yet
- tmpAB6C TMPDocument2 pagestmpAB6C TMPFrontiersNo ratings yet
- Tmp3e28 TMPDocument2 pagesTmp3e28 TMPFrontiersNo ratings yet
- Reininger 2005Document9 pagesReininger 2005willembakjesNo ratings yet
- Syphilis: Using Modern Approaches To Understand An Old DiseaseDocument9 pagesSyphilis: Using Modern Approaches To Understand An Old DiseaseAwalliantoniNo ratings yet
- Fate of Entamoeba Histolytica During Establishment of Amoebic Liver Abscess Analyzed by Quantitative Radioimaging and HistologyDocument8 pagesFate of Entamoeba Histolytica During Establishment of Amoebic Liver Abscess Analyzed by Quantitative Radioimaging and HistologyMaithreyanNandhakumarNo ratings yet
- J Immunol 2007 Rydström 5789 801Document14 pagesJ Immunol 2007 Rydström 5789 801susana1616No ratings yet
- The Immune Response To Infection With Treponema Pallidum, The Stealth PathogenDocument8 pagesThe Immune Response To Infection With Treponema Pallidum, The Stealth Pathogenhazem alzedNo ratings yet
- Host ParasiteDocument24 pagesHost ParasiteHayatul AkmaLiaNo ratings yet
- Leishmania en El MacrófagoDocument9 pagesLeishmania en El MacrófagoKatharina DPNo ratings yet
- Parasita No Intestino Do MosquitoDocument8 pagesParasita No Intestino Do MosquitoGustavo FelpeNo ratings yet
- Jir2018 6529681Document6 pagesJir2018 6529681Herry JapariNo ratings yet
- Microbial Pathogenesis: Reeta Rai, Baijnath Das, Nageshwar Choudhary, Ayantika Talukdar, Donthamsetty Nageswara RaoDocument8 pagesMicrobial Pathogenesis: Reeta Rai, Baijnath Das, Nageshwar Choudhary, Ayantika Talukdar, Donthamsetty Nageswara RaoDiego TulcanNo ratings yet
- Camp Signalling in Trypanosomatids: Role in Pathogenesis and As A Drug TargetDocument7 pagesCamp Signalling in Trypanosomatids: Role in Pathogenesis and As A Drug TargetCamilo Ernesto Araujo BarabasNo ratings yet
- Microbial Virulence Factors: Molecular SciencesDocument6 pagesMicrobial Virulence Factors: Molecular SciencesAnasNo ratings yet
- Malaria EnzimeDocument13 pagesMalaria EnzimeAnisahMahardianiNo ratings yet
- Syphilis JCI 2011 PDFDocument9 pagesSyphilis JCI 2011 PDFSteven LeonardoNo ratings yet
- Patogenia de La ToxoplasmosisDocument10 pagesPatogenia de La ToxoplasmosisSasha de la CruzNo ratings yet
- Platelets y Escherichia ColiDocument12 pagesPlatelets y Escherichia Coliedson floresNo ratings yet
- لغات التواصل الطفيلي (1) Document24 pagesلغات التواصل الطفيلي (1) Houda BouachaNo ratings yet
- Accepted Manuscript: Molecular & Biochemical ParasitologyDocument25 pagesAccepted Manuscript: Molecular & Biochemical ParasitologyColo VoltaNo ratings yet
- J 1365-2249 2002 01874 XDocument8 pagesJ 1365-2249 2002 01874 XmclimacoNo ratings yet
- Review Typhoid Fever As Cellular Microbiological Model: Rev. Inst. Med. Trop. S. Paulo 45 (4) :185-191, July-August, 2003Document7 pagesReview Typhoid Fever As Cellular Microbiological Model: Rev. Inst. Med. Trop. S. Paulo 45 (4) :185-191, July-August, 2003Tio Prima SNo ratings yet
- Quorum Sensing in African Trypanosomes: SciencedirectDocument6 pagesQuorum Sensing in African Trypanosomes: SciencedirectNithin SakthiNo ratings yet
- 2006 Cytoadhesion of Plasmodium Falciparum-Infected Erythrocytes and The Infected Placenta A Two-Way PathwayDocument12 pages2006 Cytoadhesion of Plasmodium Falciparum-Infected Erythrocytes and The Infected Placenta A Two-Way PathwaySethawud ChaikitgosiyakulNo ratings yet
- Helminth Infections - Recognition and Modulation of The Immune Response by Innate Immune CellsDocument12 pagesHelminth Infections - Recognition and Modulation of The Immune Response by Innate Immune CellsponbohacopNo ratings yet
- Pathogens 11 00811Document5 pagesPathogens 11 00811octav drutaNo ratings yet
- Immune Escape Strategies MaleriaDocument7 pagesImmune Escape Strategies MaleriaAurelia WetondieNo ratings yet
- Review Article Plasmodium Infection: Helminth Parasites Alter Protection AgainstDocument20 pagesReview Article Plasmodium Infection: Helminth Parasites Alter Protection AgainstAdrian KhomanNo ratings yet
- Parasites Evade TheDocument7 pagesParasites Evade TheMike zombieNo ratings yet
- Vaccine Amazonensis8Document16 pagesVaccine Amazonensis8Eu Meu MesmoNo ratings yet
- Jun Zou, Nathan ShankarDocument11 pagesJun Zou, Nathan ShankarHQ HQNo ratings yet
- A Critical New Pathway For Toxin Secretion?: Clinical Implications of Basic ResearchDocument2 pagesA Critical New Pathway For Toxin Secretion?: Clinical Implications of Basic Researchrahmawati aliwarmanNo ratings yet
- Diagnosis of Malaria Parasites Plasmodium Spp. in EndemicAreas: Current Strategies For An Ancient Disease Brian Gitta and Nicole KilianDocument12 pagesDiagnosis of Malaria Parasites Plasmodium Spp. in EndemicAreas: Current Strategies For An Ancient Disease Brian Gitta and Nicole KilianGitta BrianNo ratings yet
- Salmonellae Interactions With Host Process NATUREDocument15 pagesSalmonellae Interactions With Host Process NATUREDiegoNo ratings yet
- Canan 2014 JLBDocument1 pageCanan 2014 JLBNandan GokhaleNo ratings yet
- Molecular Research On Plasmodium Infection and ImmunityDocument3 pagesMolecular Research On Plasmodium Infection and ImmunityJean Bosco MBONIMPANo ratings yet
- Cimid 2015 AjayDocument7 pagesCimid 2015 AjayprashantcdriNo ratings yet
- Helmintos TisularesDocument8 pagesHelmintos TisularesValentina Arenas RNo ratings yet
- Pone 0017849Document8 pagesPone 0017849SHAFIA KHAIRANINo ratings yet
- Sap RematikDocument7 pagesSap RematikOzada RasifaNo ratings yet
- Streptococcus Group A InfectionsDocument13 pagesStreptococcus Group A InfectionsLidia GotoYourhappyplaceNo ratings yet
- Phagocytosis A Fundamental Process in ImmunityDocument19 pagesPhagocytosis A Fundamental Process in ImmunityMarco MPNo ratings yet
- CD14CD16 PDFDocument16 pagesCD14CD16 PDFAde OktiviyariNo ratings yet
- Review Article: Host Cell Signalling and Mechanisms of EvasionDocument15 pagesReview Article: Host Cell Signalling and Mechanisms of EvasionParijat BanerjeeNo ratings yet
- Bacterial Virulence Factors: Secreted For Survival: Mycobacterium TuberculosisDocument10 pagesBacterial Virulence Factors: Secreted For Survival: Mycobacterium TuberculosisRahayuNo ratings yet
- tmp8748 TMPDocument11 pagestmp8748 TMPFrontiersNo ratings yet
- tmpD859 TMPDocument7 pagestmpD859 TMPFrontiersNo ratings yet
- ThesisDocument21 pagesThesisYuti BesainNo ratings yet
- Peraza 1Document9 pagesPeraza 1api-316230227No ratings yet
- Parasitic Infections and the Immune SystemFrom EverandParasitic Infections and the Immune SystemFelipe KierzenbaumNo ratings yet
- Grade XI Chemistry EquilibriumDocument139 pagesGrade XI Chemistry EquilibriumnavyaNo ratings yet
- 3 Module Two Lesson Three Guided NotesDocument4 pages3 Module Two Lesson Three Guided NotesJoelle MatarNo ratings yet
- Do Dont - Does DoesntDocument2 pagesDo Dont - Does Doesntdeathnote lNo ratings yet
- Treatment of Constipation in Older Adult AAFPDocument8 pagesTreatment of Constipation in Older Adult AAFPTom KirklandNo ratings yet
- Revision of The Final Term Test 2022 2023Document20 pagesRevision of The Final Term Test 2022 2023Nghiêm Thị Ngọc ÁnhNo ratings yet
- Laporan Masuk N Keluar BarangDocument4 pagesLaporan Masuk N Keluar Barangmedika plazaNo ratings yet
- Count-Uncount NounsDocument2 pagesCount-Uncount NounsFrancisco Vargas100% (1)
- Maternity Benefit PolicyDocument2 pagesMaternity Benefit Policyshrija nairNo ratings yet
- Sa12mirciet9ic PDFDocument108 pagesSa12mirciet9ic PDFBobby BobbersonNo ratings yet
- NBR 14136 Plugs and Socket Outlets in BrazilDocument7 pagesNBR 14136 Plugs and Socket Outlets in BrazilCLOUD ALIGLOBALNo ratings yet
- Huawei Nova 9 Se Quick Start Guide - (Jln-lx1, Emui12.0 - 01, En-Gb, Eu)Document254 pagesHuawei Nova 9 Se Quick Start Guide - (Jln-lx1, Emui12.0 - 01, En-Gb, Eu)Dimitrije IlicNo ratings yet
- Cacanindin Mae KritikaDocument7 pagesCacanindin Mae KritikamaecacanindinNo ratings yet
- Functions of Starch and Application in Filipino DishesDocument35 pagesFunctions of Starch and Application in Filipino DishesBian Ca100% (1)
- O & M Manual - Flow Nozzle AssemblyDocument4 pagesO & M Manual - Flow Nozzle Assemblyabdellah faqdaniNo ratings yet
- Water Treatment ChemicalsDocument33 pagesWater Treatment ChemicalsMutasim Hasan MehranNo ratings yet
- Rothblum, Solomon, Murakami 1986 - Diferencias Cognitivas, Afectivas y Conductuales Entre Procrastinadores Altos y Bajos PDFDocument8 pagesRothblum, Solomon, Murakami 1986 - Diferencias Cognitivas, Afectivas y Conductuales Entre Procrastinadores Altos y Bajos PDFMartin RuízNo ratings yet
- Income Tax NotesDocument18 pagesIncome Tax NotesVikash kumarNo ratings yet
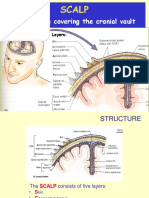
- ScalpDocument17 pagesScalpKaruna PrabhuNo ratings yet
- Safety Data Sheet: HI 7071 Reference Electrolyte For PH and Redox ElectrodesDocument3 pagesSafety Data Sheet: HI 7071 Reference Electrolyte For PH and Redox ElectrodesMagaly RojasNo ratings yet
- High Performance Plastic Materials Guide Aug 2009Document8 pagesHigh Performance Plastic Materials Guide Aug 2009JeanMichelMNo ratings yet
- LG Us-Q186c4a4 - LG Electronics LKDocument21 pagesLG Us-Q186c4a4 - LG Electronics LKAvk SanjeevanNo ratings yet
- Explosives Used in QuarryingDocument11 pagesExplosives Used in Quarryingjohn mwangiNo ratings yet
- Ammonia MeasurementDocument10 pagesAmmonia Measurement9326773777No ratings yet
- Swine ProductionDocument73 pagesSwine ProductionDUGONG PHNo ratings yet
- FadingDocument10 pagesFadingLê Dương LongNo ratings yet
- Family Business Daily: A Slice of AdviceDocument3 pagesFamily Business Daily: A Slice of Advicemaheen khanNo ratings yet
- I Am DepressedDocument8 pagesI Am DepressedwisgeorgekwokNo ratings yet