
Pi Is 0092867415013379
Pi Is 0092867415013379
You might also like
- KEMP, David. Stimulated Acoustic Emissions From Within The Human Auditory SystemDocument6 pagesKEMP, David. Stimulated Acoustic Emissions From Within The Human Auditory SystemCamanhoNo ratings yet
- ILS L4 Transcripts PDFDocument38 pagesILS L4 Transcripts PDFThuy Lan75% (4)
- Trauma Acoustic JournalDocument6 pagesTrauma Acoustic JournalLannydchandraNo ratings yet
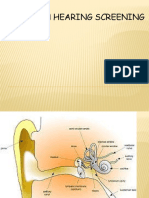
- New Born Hearing ScreeningDocument26 pagesNew Born Hearing Screeningdeadly_guyNo ratings yet
- Efficiency of Stenger Test in Confirming Profound Unilateral Pseudohypacusis PDFDocument5 pagesEfficiency of Stenger Test in Confirming Profound Unilateral Pseudohypacusis PDFValentina Muñoz OyarzúnNo ratings yet
- Bjorl: Auditory Steady State Response in Pediatric AudiologyDocument6 pagesBjorl: Auditory Steady State Response in Pediatric AudiologyNur Syamsiah MNo ratings yet
- Kakigi A Et Al - Tonotopic Mapping in Auditory Cortex of The Adult Chinchilla With Amikacin Induced Cochlear Lesions - 2000 1Document10 pagesKakigi A Et Al - Tonotopic Mapping in Auditory Cortex of The Adult Chinchilla With Amikacin Induced Cochlear Lesions - 2000 1karinduarte30No ratings yet
- Capitulo 25 PEATC en Niños e InfantesDocument27 pagesCapitulo 25 PEATC en Niños e InfantesMariela Oliva mondragonNo ratings yet
- Auditory Neuropathy Spectrum Disorders: From Diagnosis To Treatment: Literature Review and Case ReportsDocument29 pagesAuditory Neuropathy Spectrum Disorders: From Diagnosis To Treatment: Literature Review and Case Reportschipta lestariNo ratings yet
- Slow Auditory Evoked Potentials (1 InterligneDocument8 pagesSlow Auditory Evoked Potentials (1 Interligner.boniver524No ratings yet
- International Journal of Developmental Neuroscience: S.L. Uran, M.E. Gómez-Casati, L.R. GuelmanDocument11 pagesInternational Journal of Developmental Neuroscience: S.L. Uran, M.E. Gómez-Casati, L.R. GuelmanWoody WoodyNo ratings yet
- Cortical Evoked Response Audio Me TryDocument12 pagesCortical Evoked Response Audio Me TryKelvin Osamudiamen OmereNo ratings yet
- Rehab - Hi (New)Document3 pagesRehab - Hi (New)devNo ratings yet
- Prog Oftalmo 1 2019Document6 pagesProg Oftalmo 1 2019FerviferrazNo ratings yet
- Optimal Fickering Light Stimulation For Entraining Gamma Waves in The Human BrainDocument10 pagesOptimal Fickering Light Stimulation For Entraining Gamma Waves in The Human BrainPrakash N KNo ratings yet
- Acoustic and Optoacoustic Stimulations in Auditory Brainstem Response Test inDocument11 pagesAcoustic and Optoacoustic Stimulations in Auditory Brainstem Response Test inabsabravo4No ratings yet
- Hearing Disorders in Cats: Classification, Pathology and DiagnosisDocument12 pagesHearing Disorders in Cats: Classification, Pathology and Diagnosissantri safiraNo ratings yet
- Longenecker-Galazyuk2011 Article DevelopmentOfTinnitusInCBACaJMDocument12 pagesLongenecker-Galazyuk2011 Article DevelopmentOfTinnitusInCBACaJMannaNo ratings yet
- Classification of Hearing LossDocument4 pagesClassification of Hearing LossCarmen-BadeaNo ratings yet
- SorderaDocument105 pagesSorderaAzís Humberto Gastelum ZatarainNo ratings yet
- Schwannoma Vestibular TinnitusDocument6 pagesSchwannoma Vestibular TinnitusThomasMáximoMancinelliRinaldoNo ratings yet
- Effects of Noise in Animals. Turner2005Document28 pagesEffects of Noise in Animals. Turner2005Muzi Valentina RojasNo ratings yet
- Audiogram Estimation by Auditory Brainstem Response With NB CE-chirp LS Stimulus in Normal Hearing InfantsDocument7 pagesAudiogram Estimation by Auditory Brainstem Response With NB CE-chirp LS Stimulus in Normal Hearing InfantsZoe RgNo ratings yet
- Fnagi 07 00007Document14 pagesFnagi 07 00007Luana CarvalhoNo ratings yet
- 1 s2.0 S0892199721000424 MainDocument10 pages1 s2.0 S0892199721000424 Maincamilavilar2001No ratings yet
- Medicine: Characteristics of Hearing Loss in Patients With Herpes Zoster OticusDocument6 pagesMedicine: Characteristics of Hearing Loss in Patients With Herpes Zoster OticusnabilulhakNo ratings yet
- Electro-Tactile Stimulation (ETS) Enhances Cochlear-Implant Mandarin Tone RecognitionDocument5 pagesElectro-Tactile Stimulation (ETS) Enhances Cochlear-Implant Mandarin Tone RecognitionAdrian ErindraNo ratings yet
- tmp719B TMPDocument13 pagestmp719B TMPFrontiersNo ratings yet
- tmp9198 TMPDocument13 pagestmp9198 TMPFrontiersNo ratings yet
- Jurnal VertigoDocument4 pagesJurnal VertigoNur Cahyani SetiawatiNo ratings yet
- 2019 - Analisis Automatizado Del PotencialDocument8 pages2019 - Analisis Automatizado Del PotencialConny Morales BoucherNo ratings yet
- Cortical Evoked Potentials To An AuditorDocument11 pagesCortical Evoked Potentials To An Auditorjwc121.121.121No ratings yet
- Auditory Evoked Potentials - AEPs - Underlying PrinciplesDocument19 pagesAuditory Evoked Potentials - AEPs - Underlying PrinciplesMansi SinghNo ratings yet
- Disorders of AuditionDocument18 pagesDisorders of AuditionArouetNo ratings yet
- ENT NotesDocument58 pagesENT NotesJun 27No ratings yet
- Minimally Invasive Surgery For The Treatment Of.97204Document7 pagesMinimally Invasive Surgery For The Treatment Of.97204Tamilselvam KakkanaiahNo ratings yet
- Hearing Impairment in Older People: A Review: Lisa Fook, Rosemary MorganDocument5 pagesHearing Impairment in Older People: A Review: Lisa Fook, Rosemary MorganMANGKAGENo ratings yet
- J Neuron 2012 05 019Document11 pagesJ Neuron 2012 05 019Phương Nguyễn Thị ThanhNo ratings yet
- Acoustic Stapedial Rekexes in Healthy Neonates: Normative Data and Test-Retest ReliabilityDocument12 pagesAcoustic Stapedial Rekexes in Healthy Neonates: Normative Data and Test-Retest ReliabilityTeresa Eufhoria Puerto MonttNo ratings yet
- 1992 Kujala Et Al Blind ERP Aud (R)Document4 pages1992 Kujala Et Al Blind ERP Aud (R)Nidhi AggarwalNo ratings yet
- ReversedIpsilateralAcousticReflexPattern BaslDocument9 pagesReversedIpsilateralAcousticReflexPattern BaslSarah fadlainiNo ratings yet
- Classification of Hearing Loss PDFDocument4 pagesClassification of Hearing Loss PDFAsiya ZaidiNo ratings yet
- Hearing Status in Neonatal Hyperbilirubinemia by Auditory Brain Stem Evoked Response and Transient Evoked Otoacoustic EmissionDocument4 pagesHearing Status in Neonatal Hyperbilirubinemia by Auditory Brain Stem Evoked Response and Transient Evoked Otoacoustic EmissionGaurav MedikeriNo ratings yet
- Frequency Responses of Headphones Modulate Alpha Brain OscillationsDocument10 pagesFrequency Responses of Headphones Modulate Alpha Brain Oscillationsnataly Delgado MontecinosNo ratings yet
- Dies Mann 1999 StableDocument7 pagesDies Mann 1999 StablepastafarianboyNo ratings yet
- Volume - 41 - 4 - 97 - 4 - Japanese MacacaDocument7 pagesVolume - 41 - 4 - 97 - 4 - Japanese MacacaAnindyaMustikaNo ratings yet
- Otoacoustic Emissions - OAEsDocument8 pagesOtoacoustic Emissions - OAEsاحمد تحمدNo ratings yet
- Saxena 2020Document6 pagesSaxena 2020Sarah fadlainiNo ratings yet
- Toluene-Induced Hearing Loss: A Mid-Frequency Location of The Cochlear LesionsDocument12 pagesToluene-Induced Hearing Loss: A Mid-Frequency Location of The Cochlear LesionsCristian Sanabria DueñasNo ratings yet
- Tuning and Sensitivity of The Human Vestibular System To Low-Frequency VibrationDocument1 pageTuning and Sensitivity of The Human Vestibular System To Low-Frequency Vibrationsgcauri2157No ratings yet
- Overexpression of The Immediate-Early Genes Egr1, Egr2, and Egr3 in Two Strains of Rodents Susceptible To Audiogenic SeizuresDocument12 pagesOverexpression of The Immediate-Early Genes Egr1, Egr2, and Egr3 in Two Strains of Rodents Susceptible To Audiogenic SeizuresJavier HerreroNo ratings yet
- Otoacoustic Emissions From MedscapeDocument5 pagesOtoacoustic Emissions From MedscapeMin-Joo Esther ParkNo ratings yet
- Rehabilitación de PresbiciaDocument11 pagesRehabilitación de PresbiciaZuri S. GalindoNo ratings yet
- Inferior Colliculus Transcriptome After Status Epilepticus in The Genetically Audiogenic Seizure-Prone Hamster GASH/Sal Sandra M.Document19 pagesInferior Colliculus Transcriptome After Status Epilepticus in The Genetically Audiogenic Seizure-Prone Hamster GASH/Sal Sandra M.Javier HerreroNo ratings yet
- Chronic Electrical Stimulation by ADocument20 pagesChronic Electrical Stimulation by Anadir adelNo ratings yet
- Effect of Tinnitus in Distortion Products Otoacoustic Emissions (Dpoaes) in Normal Hearing PatientsDocument6 pagesEffect of Tinnitus in Distortion Products Otoacoustic Emissions (Dpoaes) in Normal Hearing Patientsvictoria barrientoNo ratings yet
- Ok Gopal Kowalski 1998 PDFDocument7 pagesOk Gopal Kowalski 1998 PDFTinha 999 chancesNo ratings yet
- MMN Reference ValuesDocument5 pagesMMN Reference ValuesLetícia Maria Martins Vasconcelos ParreiraNo ratings yet
- 1993 Alho Et Al (1993)Document10 pages1993 Alho Et Al (1993)Nidhi AggarwalNo ratings yet
- ENEURO 0396-23 2024 FullDocument44 pagesENEURO 0396-23 2024 FullAmiyangshu DeNo ratings yet
- 5) NeurosienceDocument165 pages5) NeurosienceDaBid Lopez RodriguezNo ratings yet
- Steel GuitarDocument1 pageSteel GuitarDaBid Lopez RodriguezNo ratings yet
- Steel Guitar2Document1 pageSteel Guitar2DaBid Lopez RodriguezNo ratings yet
- 21 - Taylor (2008)Document8 pages21 - Taylor (2008)DaBid Lopez RodriguezNo ratings yet
- In Psychosis, Cortical Interneurons Overexpress DNA-methyltransferase 1Document6 pagesIn Psychosis, Cortical Interneurons Overexpress DNA-methyltransferase 1DaBid Lopez RodriguezNo ratings yet
- Articulo CristianDocument5 pagesArticulo CristianDaBid Lopez RodriguezNo ratings yet
- Gozzi Et Al.,2013Document8 pagesGozzi Et Al.,2013DaBid Lopez RodriguezNo ratings yet
- Anatomy and Physiology of Salivary Gland ShivaniDocument43 pagesAnatomy and Physiology of Salivary Gland ShivaniDeepika LamichhaneNo ratings yet
- Files 2Document18 pagesFiles 2shafeeque TNo ratings yet
- MORINDA CITRIFOLIA L. (Noni) - A Review of The Scientific Validation For Its Nutritional and Therapeutic PropertiesDocument15 pagesMORINDA CITRIFOLIA L. (Noni) - A Review of The Scientific Validation For Its Nutritional and Therapeutic PropertiesDiannokaIhzaGanung100% (1)
- Water Borne DiseasesDocument16 pagesWater Borne DiseasesdrleenagowdaNo ratings yet
- C 01 Cellular Structure and FunctionDocument86 pagesC 01 Cellular Structure and FunctionCharla CNo ratings yet
- Human Platelet AntigenDocument56 pagesHuman Platelet AntigenMohandoss Murugesan100% (4)
- Otolaryngology: James A. Pfaff Gregory P. MooreDocument14 pagesOtolaryngology: James A. Pfaff Gregory P. MooreNathaniel BonardoNo ratings yet
- Growth Factors and Wound Healing 1997Document374 pagesGrowth Factors and Wound Healing 1997Vo Ha Phuong NguyenNo ratings yet
- 7 Grade Review Packet For SOLfinalDocument20 pages7 Grade Review Packet For SOLfinalabigailguashpaNo ratings yet
- Over DominanceDocument11 pagesOver DominanceJeniNo ratings yet
- BMW Rules, 2016Document13 pagesBMW Rules, 2016Pabhat KumarNo ratings yet
- IB HL BIO FULL NOTES (Onarıldı)Document874 pagesIB HL BIO FULL NOTES (Onarıldı)Sıla DenizNo ratings yet
- Amino Acid CatabolismDocument57 pagesAmino Acid CatabolismMega Nur Hesti OktaviaNo ratings yet
- PONCE - Module 4 - BSN-2 - A18Document6 pagesPONCE - Module 4 - BSN-2 - A18Ponce Kristel Mae ONo ratings yet
- 9700 w05 ErDocument19 pages9700 w05 ErwegdanNo ratings yet
- Environment and Chromosomal ChangesDocument49 pagesEnvironment and Chromosomal ChangesNiomi ButtermilkNo ratings yet
- 2017 2018 Rex Ferdinand M TraifalgarDocument63 pages2017 2018 Rex Ferdinand M Traifalgarjns1606No ratings yet
- Microbes PPT 1Document16 pagesMicrobes PPT 1BML channelNo ratings yet
- AP14 DigestiveDocument7 pagesAP14 DigestiveAmore BuenafeNo ratings yet
- Bacteriophage LambdaDocument118 pagesBacteriophage LambdaKirk SummaTime HenryNo ratings yet
- Subversion of Innate and Adaptive Immune Responses by Toxoplasma GondiiDocument13 pagesSubversion of Innate and Adaptive Immune Responses by Toxoplasma GondiiAaminnNo ratings yet
- Glomerulonefrita Acuta Poststreptococica (GNAPS) - Engleza 2 (User-PC - S Conflicted Copy 2013-04-29) - 1Document37 pagesGlomerulonefrita Acuta Poststreptococica (GNAPS) - Engleza 2 (User-PC - S Conflicted Copy 2013-04-29) - 1Ashley PaulNo ratings yet
- Gallimune 407 ND+IB+EDS+ART: PresentationDocument2 pagesGallimune 407 ND+IB+EDS+ART: PresentationRahmat SyahrirNo ratings yet
- Hematology Final ExamDocument20 pagesHematology Final ExamAlon GoldfainerNo ratings yet
- Hema OmgDocument82 pagesHema OmgBernadeth BaduaNo ratings yet
- Interaction Between Biomolecules and NanoparticlesDocument85 pagesInteraction Between Biomolecules and NanoparticlesshabanaNo ratings yet
- Application Questions and Problems Section 14.1Document8 pagesApplication Questions and Problems Section 14.1Phú NguyễnNo ratings yet
- SSLC Biology Study Material Ujwalam 2023 by DIET Kollam (Eng Med)Document79 pagesSSLC Biology Study Material Ujwalam 2023 by DIET Kollam (Eng Med)adhikesh65No ratings yet
- DNA Vaccines Prim Time Is NowDocument7 pagesDNA Vaccines Prim Time Is NowalferezhassanNo ratings yet






























































































Download as pdf or txt
You might also like
- KEMP, David. Stimulated Acoustic Emissions From Within The Human Auditory SystemDocument6 pagesKEMP, David. Stimulated Acoustic Emissions From Within The Human Auditory SystemCamanhoNo ratings yet
- ILS L4 Transcripts PDFDocument38 pagesILS L4 Transcripts PDFThuy Lan75% (4)
- Trauma Acoustic JournalDocument6 pagesTrauma Acoustic JournalLannydchandraNo ratings yet
- New Born Hearing ScreeningDocument26 pagesNew Born Hearing Screeningdeadly_guyNo ratings yet
- Efficiency of Stenger Test in Confirming Profound Unilateral Pseudohypacusis PDFDocument5 pagesEfficiency of Stenger Test in Confirming Profound Unilateral Pseudohypacusis PDFValentina Muñoz OyarzúnNo ratings yet
- Bjorl: Auditory Steady State Response in Pediatric AudiologyDocument6 pagesBjorl: Auditory Steady State Response in Pediatric AudiologyNur Syamsiah MNo ratings yet
- Kakigi A Et Al - Tonotopic Mapping in Auditory Cortex of The Adult Chinchilla With Amikacin Induced Cochlear Lesions - 2000 1Document10 pagesKakigi A Et Al - Tonotopic Mapping in Auditory Cortex of The Adult Chinchilla With Amikacin Induced Cochlear Lesions - 2000 1karinduarte30No ratings yet
- Capitulo 25 PEATC en Niños e InfantesDocument27 pagesCapitulo 25 PEATC en Niños e InfantesMariela Oliva mondragonNo ratings yet
- Auditory Neuropathy Spectrum Disorders: From Diagnosis To Treatment: Literature Review and Case ReportsDocument29 pagesAuditory Neuropathy Spectrum Disorders: From Diagnosis To Treatment: Literature Review and Case Reportschipta lestariNo ratings yet
- Slow Auditory Evoked Potentials (1 InterligneDocument8 pagesSlow Auditory Evoked Potentials (1 Interligner.boniver524No ratings yet
- International Journal of Developmental Neuroscience: S.L. Uran, M.E. Gómez-Casati, L.R. GuelmanDocument11 pagesInternational Journal of Developmental Neuroscience: S.L. Uran, M.E. Gómez-Casati, L.R. GuelmanWoody WoodyNo ratings yet
- Cortical Evoked Response Audio Me TryDocument12 pagesCortical Evoked Response Audio Me TryKelvin Osamudiamen OmereNo ratings yet
- Rehab - Hi (New)Document3 pagesRehab - Hi (New)devNo ratings yet
- Prog Oftalmo 1 2019Document6 pagesProg Oftalmo 1 2019FerviferrazNo ratings yet
- Optimal Fickering Light Stimulation For Entraining Gamma Waves in The Human BrainDocument10 pagesOptimal Fickering Light Stimulation For Entraining Gamma Waves in The Human BrainPrakash N KNo ratings yet
- Acoustic and Optoacoustic Stimulations in Auditory Brainstem Response Test inDocument11 pagesAcoustic and Optoacoustic Stimulations in Auditory Brainstem Response Test inabsabravo4No ratings yet
- Hearing Disorders in Cats: Classification, Pathology and DiagnosisDocument12 pagesHearing Disorders in Cats: Classification, Pathology and Diagnosissantri safiraNo ratings yet
- Longenecker-Galazyuk2011 Article DevelopmentOfTinnitusInCBACaJMDocument12 pagesLongenecker-Galazyuk2011 Article DevelopmentOfTinnitusInCBACaJMannaNo ratings yet
- Classification of Hearing LossDocument4 pagesClassification of Hearing LossCarmen-BadeaNo ratings yet
- SorderaDocument105 pagesSorderaAzís Humberto Gastelum ZatarainNo ratings yet
- Schwannoma Vestibular TinnitusDocument6 pagesSchwannoma Vestibular TinnitusThomasMáximoMancinelliRinaldoNo ratings yet
- Effects of Noise in Animals. Turner2005Document28 pagesEffects of Noise in Animals. Turner2005Muzi Valentina RojasNo ratings yet
- Audiogram Estimation by Auditory Brainstem Response With NB CE-chirp LS Stimulus in Normal Hearing InfantsDocument7 pagesAudiogram Estimation by Auditory Brainstem Response With NB CE-chirp LS Stimulus in Normal Hearing InfantsZoe RgNo ratings yet
- Fnagi 07 00007Document14 pagesFnagi 07 00007Luana CarvalhoNo ratings yet
- 1 s2.0 S0892199721000424 MainDocument10 pages1 s2.0 S0892199721000424 Maincamilavilar2001No ratings yet
- Medicine: Characteristics of Hearing Loss in Patients With Herpes Zoster OticusDocument6 pagesMedicine: Characteristics of Hearing Loss in Patients With Herpes Zoster OticusnabilulhakNo ratings yet
- Electro-Tactile Stimulation (ETS) Enhances Cochlear-Implant Mandarin Tone RecognitionDocument5 pagesElectro-Tactile Stimulation (ETS) Enhances Cochlear-Implant Mandarin Tone RecognitionAdrian ErindraNo ratings yet
- tmp719B TMPDocument13 pagestmp719B TMPFrontiersNo ratings yet
- tmp9198 TMPDocument13 pagestmp9198 TMPFrontiersNo ratings yet
- Jurnal VertigoDocument4 pagesJurnal VertigoNur Cahyani SetiawatiNo ratings yet
- 2019 - Analisis Automatizado Del PotencialDocument8 pages2019 - Analisis Automatizado Del PotencialConny Morales BoucherNo ratings yet
- Cortical Evoked Potentials To An AuditorDocument11 pagesCortical Evoked Potentials To An Auditorjwc121.121.121No ratings yet
- Auditory Evoked Potentials - AEPs - Underlying PrinciplesDocument19 pagesAuditory Evoked Potentials - AEPs - Underlying PrinciplesMansi SinghNo ratings yet
- Disorders of AuditionDocument18 pagesDisorders of AuditionArouetNo ratings yet
- ENT NotesDocument58 pagesENT NotesJun 27No ratings yet
- Minimally Invasive Surgery For The Treatment Of.97204Document7 pagesMinimally Invasive Surgery For The Treatment Of.97204Tamilselvam KakkanaiahNo ratings yet
- Hearing Impairment in Older People: A Review: Lisa Fook, Rosemary MorganDocument5 pagesHearing Impairment in Older People: A Review: Lisa Fook, Rosemary MorganMANGKAGENo ratings yet
- J Neuron 2012 05 019Document11 pagesJ Neuron 2012 05 019Phương Nguyễn Thị ThanhNo ratings yet
- Acoustic Stapedial Rekexes in Healthy Neonates: Normative Data and Test-Retest ReliabilityDocument12 pagesAcoustic Stapedial Rekexes in Healthy Neonates: Normative Data and Test-Retest ReliabilityTeresa Eufhoria Puerto MonttNo ratings yet
- 1992 Kujala Et Al Blind ERP Aud (R)Document4 pages1992 Kujala Et Al Blind ERP Aud (R)Nidhi AggarwalNo ratings yet
- ReversedIpsilateralAcousticReflexPattern BaslDocument9 pagesReversedIpsilateralAcousticReflexPattern BaslSarah fadlainiNo ratings yet
- Classification of Hearing Loss PDFDocument4 pagesClassification of Hearing Loss PDFAsiya ZaidiNo ratings yet
- Hearing Status in Neonatal Hyperbilirubinemia by Auditory Brain Stem Evoked Response and Transient Evoked Otoacoustic EmissionDocument4 pagesHearing Status in Neonatal Hyperbilirubinemia by Auditory Brain Stem Evoked Response and Transient Evoked Otoacoustic EmissionGaurav MedikeriNo ratings yet
- Frequency Responses of Headphones Modulate Alpha Brain OscillationsDocument10 pagesFrequency Responses of Headphones Modulate Alpha Brain Oscillationsnataly Delgado MontecinosNo ratings yet
- Dies Mann 1999 StableDocument7 pagesDies Mann 1999 StablepastafarianboyNo ratings yet
- Volume - 41 - 4 - 97 - 4 - Japanese MacacaDocument7 pagesVolume - 41 - 4 - 97 - 4 - Japanese MacacaAnindyaMustikaNo ratings yet
- Otoacoustic Emissions - OAEsDocument8 pagesOtoacoustic Emissions - OAEsاحمد تحمدNo ratings yet
- Saxena 2020Document6 pagesSaxena 2020Sarah fadlainiNo ratings yet
- Toluene-Induced Hearing Loss: A Mid-Frequency Location of The Cochlear LesionsDocument12 pagesToluene-Induced Hearing Loss: A Mid-Frequency Location of The Cochlear LesionsCristian Sanabria DueñasNo ratings yet
- Tuning and Sensitivity of The Human Vestibular System To Low-Frequency VibrationDocument1 pageTuning and Sensitivity of The Human Vestibular System To Low-Frequency Vibrationsgcauri2157No ratings yet
- Overexpression of The Immediate-Early Genes Egr1, Egr2, and Egr3 in Two Strains of Rodents Susceptible To Audiogenic SeizuresDocument12 pagesOverexpression of The Immediate-Early Genes Egr1, Egr2, and Egr3 in Two Strains of Rodents Susceptible To Audiogenic SeizuresJavier HerreroNo ratings yet
- Otoacoustic Emissions From MedscapeDocument5 pagesOtoacoustic Emissions From MedscapeMin-Joo Esther ParkNo ratings yet
- Rehabilitación de PresbiciaDocument11 pagesRehabilitación de PresbiciaZuri S. GalindoNo ratings yet
- Inferior Colliculus Transcriptome After Status Epilepticus in The Genetically Audiogenic Seizure-Prone Hamster GASH/Sal Sandra M.Document19 pagesInferior Colliculus Transcriptome After Status Epilepticus in The Genetically Audiogenic Seizure-Prone Hamster GASH/Sal Sandra M.Javier HerreroNo ratings yet
- Chronic Electrical Stimulation by ADocument20 pagesChronic Electrical Stimulation by Anadir adelNo ratings yet
- Effect of Tinnitus in Distortion Products Otoacoustic Emissions (Dpoaes) in Normal Hearing PatientsDocument6 pagesEffect of Tinnitus in Distortion Products Otoacoustic Emissions (Dpoaes) in Normal Hearing Patientsvictoria barrientoNo ratings yet
- Ok Gopal Kowalski 1998 PDFDocument7 pagesOk Gopal Kowalski 1998 PDFTinha 999 chancesNo ratings yet
- MMN Reference ValuesDocument5 pagesMMN Reference ValuesLetícia Maria Martins Vasconcelos ParreiraNo ratings yet
- 1993 Alho Et Al (1993)Document10 pages1993 Alho Et Al (1993)Nidhi AggarwalNo ratings yet
- ENEURO 0396-23 2024 FullDocument44 pagesENEURO 0396-23 2024 FullAmiyangshu DeNo ratings yet
- 5) NeurosienceDocument165 pages5) NeurosienceDaBid Lopez RodriguezNo ratings yet
- Steel GuitarDocument1 pageSteel GuitarDaBid Lopez RodriguezNo ratings yet
- Steel Guitar2Document1 pageSteel Guitar2DaBid Lopez RodriguezNo ratings yet
- 21 - Taylor (2008)Document8 pages21 - Taylor (2008)DaBid Lopez RodriguezNo ratings yet
- In Psychosis, Cortical Interneurons Overexpress DNA-methyltransferase 1Document6 pagesIn Psychosis, Cortical Interneurons Overexpress DNA-methyltransferase 1DaBid Lopez RodriguezNo ratings yet
- Articulo CristianDocument5 pagesArticulo CristianDaBid Lopez RodriguezNo ratings yet
- Gozzi Et Al.,2013Document8 pagesGozzi Et Al.,2013DaBid Lopez RodriguezNo ratings yet
- Anatomy and Physiology of Salivary Gland ShivaniDocument43 pagesAnatomy and Physiology of Salivary Gland ShivaniDeepika LamichhaneNo ratings yet
- Files 2Document18 pagesFiles 2shafeeque TNo ratings yet
- MORINDA CITRIFOLIA L. (Noni) - A Review of The Scientific Validation For Its Nutritional and Therapeutic PropertiesDocument15 pagesMORINDA CITRIFOLIA L. (Noni) - A Review of The Scientific Validation For Its Nutritional and Therapeutic PropertiesDiannokaIhzaGanung100% (1)
- Water Borne DiseasesDocument16 pagesWater Borne DiseasesdrleenagowdaNo ratings yet
- C 01 Cellular Structure and FunctionDocument86 pagesC 01 Cellular Structure and FunctionCharla CNo ratings yet
- Human Platelet AntigenDocument56 pagesHuman Platelet AntigenMohandoss Murugesan100% (4)
- Otolaryngology: James A. Pfaff Gregory P. MooreDocument14 pagesOtolaryngology: James A. Pfaff Gregory P. MooreNathaniel BonardoNo ratings yet
- Growth Factors and Wound Healing 1997Document374 pagesGrowth Factors and Wound Healing 1997Vo Ha Phuong NguyenNo ratings yet
- 7 Grade Review Packet For SOLfinalDocument20 pages7 Grade Review Packet For SOLfinalabigailguashpaNo ratings yet
- Over DominanceDocument11 pagesOver DominanceJeniNo ratings yet
- BMW Rules, 2016Document13 pagesBMW Rules, 2016Pabhat KumarNo ratings yet
- IB HL BIO FULL NOTES (Onarıldı)Document874 pagesIB HL BIO FULL NOTES (Onarıldı)Sıla DenizNo ratings yet
- Amino Acid CatabolismDocument57 pagesAmino Acid CatabolismMega Nur Hesti OktaviaNo ratings yet
- PONCE - Module 4 - BSN-2 - A18Document6 pagesPONCE - Module 4 - BSN-2 - A18Ponce Kristel Mae ONo ratings yet
- 9700 w05 ErDocument19 pages9700 w05 ErwegdanNo ratings yet
- Environment and Chromosomal ChangesDocument49 pagesEnvironment and Chromosomal ChangesNiomi ButtermilkNo ratings yet
- 2017 2018 Rex Ferdinand M TraifalgarDocument63 pages2017 2018 Rex Ferdinand M Traifalgarjns1606No ratings yet
- Microbes PPT 1Document16 pagesMicrobes PPT 1BML channelNo ratings yet
- AP14 DigestiveDocument7 pagesAP14 DigestiveAmore BuenafeNo ratings yet
- Bacteriophage LambdaDocument118 pagesBacteriophage LambdaKirk SummaTime HenryNo ratings yet
- Subversion of Innate and Adaptive Immune Responses by Toxoplasma GondiiDocument13 pagesSubversion of Innate and Adaptive Immune Responses by Toxoplasma GondiiAaminnNo ratings yet
- Glomerulonefrita Acuta Poststreptococica (GNAPS) - Engleza 2 (User-PC - S Conflicted Copy 2013-04-29) - 1Document37 pagesGlomerulonefrita Acuta Poststreptococica (GNAPS) - Engleza 2 (User-PC - S Conflicted Copy 2013-04-29) - 1Ashley PaulNo ratings yet
- Gallimune 407 ND+IB+EDS+ART: PresentationDocument2 pagesGallimune 407 ND+IB+EDS+ART: PresentationRahmat SyahrirNo ratings yet
- Hematology Final ExamDocument20 pagesHematology Final ExamAlon GoldfainerNo ratings yet
- Hema OmgDocument82 pagesHema OmgBernadeth BaduaNo ratings yet
- Interaction Between Biomolecules and NanoparticlesDocument85 pagesInteraction Between Biomolecules and NanoparticlesshabanaNo ratings yet
- Application Questions and Problems Section 14.1Document8 pagesApplication Questions and Problems Section 14.1Phú NguyễnNo ratings yet
- SSLC Biology Study Material Ujwalam 2023 by DIET Kollam (Eng Med)Document79 pagesSSLC Biology Study Material Ujwalam 2023 by DIET Kollam (Eng Med)adhikesh65No ratings yet
- DNA Vaccines Prim Time Is NowDocument7 pagesDNA Vaccines Prim Time Is NowalferezhassanNo ratings yet