Download as pdf or txt
You might also like
- PD 705 Revised Forestry Code of The PhilippinesDocument52 pagesPD 705 Revised Forestry Code of The PhilippinesTine Tungpalan83% (6)
- Grande Island, Subic Zambales Seagrass Monitoring 2023 (1-03-24)Document14 pagesGrande Island, Subic Zambales Seagrass Monitoring 2023 (1-03-24)Clinton BernalesNo ratings yet
- Jellyfish Aggregations and Leatherback Turtle Foraging Patterns in A Temperate Coastal EnvironmentDocument6 pagesJellyfish Aggregations and Leatherback Turtle Foraging Patterns in A Temperate Coastal EnvironmentBengt HörbergNo ratings yet
- This Is The Published VersionDocument7 pagesThis Is The Published VersionAlínio JoséNo ratings yet
- Griffithsia Capitata 2016Document5 pagesGriffithsia Capitata 2016PAOLA fRANCONo ratings yet
- Occurrence, Distribution and Composition of Marine Mammals in The Bight of Bony, NigeriaDocument10 pagesOccurrence, Distribution and Composition of Marine Mammals in The Bight of Bony, NigeriaRizky RamaNo ratings yet
- Theory of Island Biogeography Lab ActivityDocument2 pagesTheory of Island Biogeography Lab ActivityChances chanNo ratings yet
- Carranza 2006 BiolConservDocument6 pagesCarranza 2006 BiolConservtortugamarina100% (2)
- 13 Esponjas Porifera Parque Nacional San Esteban 2010Document14 pages13 Esponjas Porifera Parque Nacional San Esteban 2010carmenNo ratings yet
- 16.2.4 Why Systems Respond Differently To Predator RemovalDocument4 pages16.2.4 Why Systems Respond Differently To Predator RemovalMuhammad AfifudinNo ratings yet
- JURNAL Amkasmaja UASsdDocument12 pagesJURNAL Amkasmaja UASsdFANI INDAHSARINo ratings yet
- Survival of Juvenile Caribbean Spiny Lobster: Effects of Shelter Size, Geographic Location andDocument12 pagesSurvival of Juvenile Caribbean Spiny Lobster: Effects of Shelter Size, Geographic Location andAnonymous EoivOpNo ratings yet
- Diamond 1972 - 1Document5 pagesDiamond 1972 - 1Ade KurniawanNo ratings yet
- Egan 2004Document13 pagesEgan 2004RAKA RYANDRANo ratings yet
- Coral Community Structure at Isla Lobos Reef, Gulf of MexicoDocument4 pagesCoral Community Structure at Isla Lobos Reef, Gulf of MexicoRafael Junnar DumalanNo ratings yet
- Giglio 2017Document7 pagesGiglio 2017Yuyu FamsNo ratings yet
- Diet Waterbirds 28 106-109 1Document4 pagesDiet Waterbirds 28 106-109 1abdhamzaNo ratings yet
- Prioritizing The World's Islands For Vertebrate-Eradication ProgrammesDocument11 pagesPrioritizing The World's Islands For Vertebrate-Eradication ProgrammesThais Martins ヅNo ratings yet
- Small Cetacean Captures in Peruvian Artisanal Fisheries. High Despite. Mangel, Jeffrey C. Alfaro Shigueto, Joanna-2009Document8 pagesSmall Cetacean Captures in Peruvian Artisanal Fisheries. High Despite. Mangel, Jeffrey C. Alfaro Shigueto, Joanna-2009Yomira Leon Santa CruzNo ratings yet
- 1-s2.0-S168742851400003X (MMI) 5Document8 pages1-s2.0-S168742851400003X (MMI) 5Agus RomadhonNo ratings yet
- Layla Hollingsworth - The Island Biogeography Simulator - 14405554Document7 pagesLayla Hollingsworth - The Island Biogeography Simulator - 14405554llhllngswrthNo ratings yet
- Moncada 1999 ChelConservBiolDocument7 pagesMoncada 1999 ChelConservBiolThe Science Exchange International Sea Turtle InternshipsNo ratings yet
- Gollinetal 2021 NZJEDocument10 pagesGollinetal 2021 NZJEEtienne OssonaNo ratings yet
- Alr 96108Document7 pagesAlr 96108Trade TechnologiesNo ratings yet
- American Society of Ichthyologists and Herpetologists (ASIH), Allen Press CopeiaDocument5 pagesAmerican Society of Ichthyologists and Herpetologists (ASIH), Allen Press CopeiaDébora Samira G. NegrãoNo ratings yet
- Adaptive Radiation Versus Intraspeci®c Differentiation: Morphological Variation in Caribbean Anolis LizardsDocument6 pagesAdaptive Radiation Versus Intraspeci®c Differentiation: Morphological Variation in Caribbean Anolis LizardsMicaela GonellaNo ratings yet
- AMRE2417Document9 pagesAMRE2417tortugamarinaNo ratings yet
- Distribution and Nesting Density of The Philippine Eagle Pithecophaga Jefferyi On Mindanao Island, Philippines: What Do We Know After 100 Years?Document6 pagesDistribution and Nesting Density of The Philippine Eagle Pithecophaga Jefferyi On Mindanao Island, Philippines: What Do We Know After 100 Years?John Jerald VillamancaNo ratings yet
- Ecological Estudies of Fish Fauna Associated With M. Pyrifera in Fueguian Aislands (Moreno y Jara, 1984)Document9 pagesEcological Estudies of Fish Fauna Associated With M. Pyrifera in Fueguian Aislands (Moreno y Jara, 1984)Francisco J. OvalleNo ratings yet
- Ecology of WinteringDocument12 pagesEcology of Winteringbiologia007No ratings yet
- Cayman Islands National Biodiversity Action Plan 2009 3.T.2.1 Terrestrial Species - Invertebrates Little Cayman SnailDocument4 pagesCayman Islands National Biodiversity Action Plan 2009 3.T.2.1 Terrestrial Species - Invertebrates Little Cayman Snailsnaip1370No ratings yet
- Del Norte - Campos - A Survey of Macro-Invertebrate GleaningDocument10 pagesDel Norte - Campos - A Survey of Macro-Invertebrate GleaningfurkonableNo ratings yet
- WIF26 17 SinghDocument7 pagesWIF26 17 SinghMaybel BalansagNo ratings yet
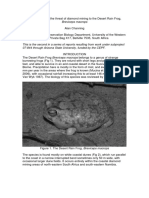
- Desert Rain Frog ThreatsDocument10 pagesDesert Rain Frog ThreatsKinokoNo ratings yet
- On The Coast of Pulau Banyak, Indonesia: October 1999Document19 pagesOn The Coast of Pulau Banyak, Indonesia: October 1999Monica KuhonNo ratings yet
- Estructuras de Las Comunidades de Corales y Octocoral de Isla de AvesDocument23 pagesEstructuras de Las Comunidades de Corales y Octocoral de Isla de AvesAtahualpaNo ratings yet
- Do Partial Marine ReserveDocument11 pagesDo Partial Marine ReserveSuper DikyNo ratings yet
- 2020 - Hernandez Landa Et Al - Size-Frequency Distribution of CoralDocument30 pages2020 - Hernandez Landa Et Al - Size-Frequency Distribution of CoralRoberto Hernandez-landaNo ratings yet
- Ecology of Antarctic Marine Sponges: An OverviewDocument10 pagesEcology of Antarctic Marine Sponges: An Overviewvltg_35643No ratings yet
- Norwood Island Mussel Study Report 2017Document21 pagesNorwood Island Mussel Study Report 2017Travis Williams100% (1)
- Is Black Swan Grazing A Threat To Seagrass? Indications From An Observational Study in New ZealandDocument10 pagesIs Black Swan Grazing A Threat To Seagrass? Indications From An Observational Study in New ZealandPrincy CaesarNo ratings yet
- Quantifying Tidal Movements of The Shore Crab Carcinus Maenas On To Complex Epibenthic Bivalve HabitatsDocument15 pagesQuantifying Tidal Movements of The Shore Crab Carcinus Maenas On To Complex Epibenthic Bivalve HabitatsbambilovespascalNo ratings yet
- Horrocks - Scott - Barbados - 1991Document8 pagesHorrocks - Scott - Barbados - 1991The Science Exchange International Sea Turtle InternshipsNo ratings yet
- Navarro Et Al, 2005Document7 pagesNavarro Et Al, 2005Carlos MeirellesNo ratings yet
- 2019-Assessment On Recruitment Density of Branching Corals Montipora Digitata (Scleractinia Acroporidae) in Talairi Island, Gulf of MannarDocument4 pages2019-Assessment On Recruitment Density of Branching Corals Montipora Digitata (Scleractinia Acroporidae) in Talairi Island, Gulf of MannarRameshNo ratings yet
- The Impact On Native Alaskan Wildlife Populations Following The Exxon Valdez Oil SpillDocument4 pagesThe Impact On Native Alaskan Wildlife Populations Following The Exxon Valdez Oil Spillapi-516918481No ratings yet
- 2019 Anthropogenic Debris Accumulated in Nests of Seabirds in An Uninhabited Island in West AfricaDocument7 pages2019 Anthropogenic Debris Accumulated in Nests of Seabirds in An Uninhabited Island in West AfricaVictoria Estrella CanulNo ratings yet
- Identifying Spatial and Temporal Patterns of Lobster Abundances in The San Andres Archipelago: An Experience of Co-ManagementDocument11 pagesIdentifying Spatial and Temporal Patterns of Lobster Abundances in The San Andres Archipelago: An Experience of Co-Managementalexpc1988No ratings yet
- Developing Bird-Strike Risk Assessment Models For Open-Water RestDocument13 pagesDeveloping Bird-Strike Risk Assessment Models For Open-Water RestBaramee MeeboonNo ratings yet
- Waluda Et Al., 2006. Influence of Enso in D.gigas Fishery in PeruDocument8 pagesWaluda Et Al., 2006. Influence of Enso in D.gigas Fishery in PeruEdward F Ccasani de la CruzNo ratings yet
- Re-Evaluation of The Extent of Caulerpa Taxifolia Development in The Northern Mediterranean Using Airborne Spectrographic SensingDocument8 pagesRe-Evaluation of The Extent of Caulerpa Taxifolia Development in The Northern Mediterranean Using Airborne Spectrographic SensingjohnNo ratings yet
- Coral in Healthy PopulationDocument12 pagesCoral in Healthy PopulationfaizzaneNo ratings yet
- Arthropods Community of Mangrove Swamp of Great Kwa River, Southern NigeriaDocument6 pagesArthropods Community of Mangrove Swamp of Great Kwa River, Southern NigeriafebyNo ratings yet
- The Impacts of Aquacultured Oysters, Crassostrea Virginica (Gmelin, 1791) On Water Column Nitrogen and Sedimentation: Results of A Mesocosm StudyDocument16 pagesThe Impacts of Aquacultured Oysters, Crassostrea Virginica (Gmelin, 1791) On Water Column Nitrogen and Sedimentation: Results of A Mesocosm Studyonceuponadream504No ratings yet
- Fmars 08 617921Document16 pagesFmars 08 617921mark españolaNo ratings yet
- Behavioural Interactions of Seabirds With Suspended Mussel LonglinesDocument12 pagesBehavioural Interactions of Seabirds With Suspended Mussel LonglinesJorge RodriguezNo ratings yet
- Ganeetal2021.BeachprofilingandghostcrabdensitiesDocument10 pagesGaneetal2021.BeachprofilingandghostcrabdensitiesMarco De GuzmanNo ratings yet
- Boa Constrictor, An Introduced Predator On Cozumel Island, MexicoDocument7 pagesBoa Constrictor, An Introduced Predator On Cozumel Island, MexicoCristopher GonzalezNo ratings yet
- 1997 RobertsIrving Bell JAridEnvirDocument9 pages1997 RobertsIrving Bell JAridEnvirrm6.gfatmNo ratings yet
- Ejemplo de Artículo Cientifico PDFDocument9 pagesEjemplo de Artículo Cientifico PDFetnoeducacionNo ratings yet
- HCV-Screening-Guidance Final May27 2019Document46 pagesHCV-Screening-Guidance Final May27 2019Gestión Documental BioApNo ratings yet
- On-Line Activity: Environmentalism Anti-EnvironmentalismDocument4 pagesOn-Line Activity: Environmentalism Anti-EnvironmentalismDaniel TabordaNo ratings yet
- 2024-Cv - AnglaisDocument1 page2024-Cv - AnglaisPriscilaBeléziaNo ratings yet
- The Influence of Riparian Vegetation and Season On Stream Metabolism of Valley Creek, MinnesotaDocument21 pagesThe Influence of Riparian Vegetation and Season On Stream Metabolism of Valley Creek, Minnesotajherrera32No ratings yet
- This Study Resource Was: Week 3Document7 pagesThis Study Resource Was: Week 3Rica Paola G. AquinoNo ratings yet
- Biblio Himalaya FloraDocument280 pagesBiblio Himalaya FloraMadhan RajNo ratings yet
- Azolla & SRI Information LeafletDocument2 pagesAzolla & SRI Information LeafletSubha Devi0% (1)
- Biodiversity and The Healthy Society: Amazon RainforestDocument16 pagesBiodiversity and The Healthy Society: Amazon RainforestEzekiel InfantadoNo ratings yet
- Overfishing Multimedia ScriptDocument7 pagesOverfishing Multimedia Scriptapi-349251913No ratings yet
- HT WEEK 6 Heritage SitesDocument4 pagesHT WEEK 6 Heritage SitesNovel LampitocNo ratings yet
- Pestle PorterDocument4 pagesPestle PorterIlham RNo ratings yet
- Deforestation in Obubra Local Government Area The ChallengesDocument7 pagesDeforestation in Obubra Local Government Area The ChallengesAlexander Decker100% (1)
- Fisheries CodeDocument34 pagesFisheries CodeMark Andrei GubacNo ratings yet
- Ecological Issues and Business DevelopmentDocument16 pagesEcological Issues and Business DevelopmentМаша ЛегуцкаяNo ratings yet
- Diversity: The Indian Sundarban Mangrove Forests: History, Utilization, Conservation Strategies and Local PerceptionDocument21 pagesDiversity: The Indian Sundarban Mangrove Forests: History, Utilization, Conservation Strategies and Local PerceptionSanghita BhowmickNo ratings yet
- Eco FarmingDocument3 pagesEco FarmingiffatNo ratings yet
- 4.1 Species, Communities and Ecosystems PracticeDocument4 pages4.1 Species, Communities and Ecosystems PracticeJames ArpinNo ratings yet
- Module 3Document7 pagesModule 3Rana VergaraNo ratings yet
- Patumahoe Hill Community Mound - Summit With A View - ForeverDocument32 pagesPatumahoe Hill Community Mound - Summit With A View - ForeverPatumahoeVillageIncNo ratings yet
- National Policy On Peatlands Management in Indonesia-SPM-Pojok IKLIM-27042018-MinDocument34 pagesNational Policy On Peatlands Management in Indonesia-SPM-Pojok IKLIM-27042018-MinDLH Kab. FlotimNo ratings yet
- Marine Biology Literature ReviewDocument4 pagesMarine Biology Literature Reviewlqinlccnd100% (1)
- Cdi 10 Compilation of 18 Environmental LawsDocument123 pagesCdi 10 Compilation of 18 Environmental LawsMa Francesca Chanelle AguilusNo ratings yet
- Ge3451-Environmental Sciencesand Sustainability-753946292-Ge3451-Environmental Sciences and Sustainability QBDocument8 pagesGe3451-Environmental Sciencesand Sustainability-753946292-Ge3451-Environmental Sciences and Sustainability QBGururaj SivaprakasamNo ratings yet
- Forest ManagementDocument9 pagesForest ManagementLynn CrastoNo ratings yet
- Unit 12: Sustainable DevelopmentDocument9 pagesUnit 12: Sustainable DevelopmentJyotsna MahawanNo ratings yet
- Bandora Panchayat - Draft - 3 - SLC Final 1 ColourDocument1 pageBandora Panchayat - Draft - 3 - SLC Final 1 ColourBhuvana EswariNo ratings yet
- Nature Based Solutions For Conservation and Management of Ullal Lake, Bangalore, Karnataka, IndiaDocument10 pagesNature Based Solutions For Conservation and Management of Ullal Lake, Bangalore, Karnataka, IndiaInternational Journal of Innovative Science and Research Technology100% (1)
- Unit-11 Global InitiativesDocument17 pagesUnit-11 Global Initiativeswinchester2000No ratings yet
- Energy Cycle: Environmental Studies. Janhavi Dalvi. ROLL NO:1914Document14 pagesEnergy Cycle: Environmental Studies. Janhavi Dalvi. ROLL NO:1914Zoya ShaikhNo ratings yet