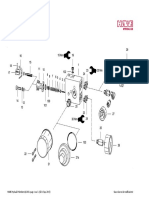
Download as pdf or txt
You might also like
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5820)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1093)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (845)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (590)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (898)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (540)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (349)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (822)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (122)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (401)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (74)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- Heights (Working At) : S W M S (SWMS) P 1Document14 pagesHeights (Working At) : S W M S (SWMS) P 1Benouna Fert100% (1)
- A Smart Motor Controller For E-Bike Applications PDFDocument4 pagesA Smart Motor Controller For E-Bike Applications PDFaungwinnaing100% (1)
- Denman - On FortificationDocument17 pagesDenman - On FortificationkontarexNo ratings yet
- Six C's of Effective MessagesDocument40 pagesSix C's of Effective MessagessheilaNo ratings yet
- Sccan Resourcemanual Allpages Update v2Document154 pagesSccan Resourcemanual Allpages Update v2SiangNo ratings yet
- The Duality of Human Nature in Oscar Wilde's The Importance of Being EarnestDocument28 pagesThe Duality of Human Nature in Oscar Wilde's The Importance of Being EarnestSowmya ShreeNo ratings yet
- Irda Jun10Document96 pagesIrda Jun10ManiNo ratings yet
- Federalism: Teaching MaterialDocument514 pagesFederalism: Teaching MaterialBirhane SeidNo ratings yet
- Invoice: PT - Sitc IndonesiaDocument1 pageInvoice: PT - Sitc IndonesiaMuhammad SyukurNo ratings yet
- Final DKSSKDocument79 pagesFinal DKSSKSarita Latthe100% (2)
- Tender InformationDocument167 pagesTender InformationComments ModeratorNo ratings yet
- Department of Education: Republic of The PhilippinesDocument4 pagesDepartment of Education: Republic of The PhilippinesLOIDA AGUILARNo ratings yet
- D 12332Document108 pagesD 12332jesus_hfNo ratings yet
- Arizona's Top Kitchen & Bath Remodeling ContractorDocument8 pagesArizona's Top Kitchen & Bath Remodeling ContractorPremier Kitchen and BathNo ratings yet
- RAMMINGDocument3 pagesRAMMINGmatheswaran a sNo ratings yet
- Chapter 2 Indian Economy 1950-1990Document27 pagesChapter 2 Indian Economy 1950-1990Ajay pandeyNo ratings yet
- Electrical Specification PDFDocument235 pagesElectrical Specification PDFMinhTrieu100% (1)
- ECE4740: Digital VLSI Design: Semiconductor MemoriesDocument35 pagesECE4740: Digital VLSI Design: Semiconductor Memoriessenthil kumar ganesanNo ratings yet
- A1 (2 3 4) F./... D 6905 Af/1Document3 pagesA1 (2 3 4) F./... D 6905 Af/1GERALD SIMONNo ratings yet
- Asb - 212 10 140Document12 pagesAsb - 212 10 140smartasss100% (1)
- The Autism Diagnostic Observation Schedule, Module 4: Application of The Revised Algorithms in An Independent, Well-Defined, Dutch Sample (N 93)Document11 pagesThe Autism Diagnostic Observation Schedule, Module 4: Application of The Revised Algorithms in An Independent, Well-Defined, Dutch Sample (N 93)Laura CamusNo ratings yet
- CBC TMDocument108 pagesCBC TMChryz SantosNo ratings yet
- Contemporary Philippine Arts From The Regions Grade 12 ModuleDocument25 pagesContemporary Philippine Arts From The Regions Grade 12 ModuleFaye Corazon GaurinoNo ratings yet
- Book Reviews: Supplemental Damping and Seismic Isolation by C. ChristopoulosDocument1 pageBook Reviews: Supplemental Damping and Seismic Isolation by C. ChristopoulosLuca AcetoNo ratings yet
- Pharmacognosy-Large FontDocument35 pagesPharmacognosy-Large FontArantxa HilarioNo ratings yet
- Extracting DnaDocument6 pagesExtracting DnaIya AlabastroNo ratings yet
- Vitamin A & D: Chemistry, Biochemical Role, RDA & Deficiency DiseasesDocument20 pagesVitamin A & D: Chemistry, Biochemical Role, RDA & Deficiency DiseasesSELVI ANo ratings yet
- NO Memo No. 21 S. 2018 Adherence To Training Policy PDFDocument10 pagesNO Memo No. 21 S. 2018 Adherence To Training Policy PDFKemberly Semaña PentonNo ratings yet
- Skript Version30 PDFDocument90 pagesSkript Version30 PDFNguyễn TúNo ratings yet
- (THAAD) Five Failures and CountingDocument6 pages(THAAD) Five Failures and CountingGuilherme Da Silva CostaNo ratings yet