Download as pdf or txt
You might also like
- Cycle SyncingDocument43 pagesCycle SyncingHeShot MeDown100% (2)
- UPSC New Syllabus & Tips To Crack IAS Preliminary & Mains Exam PDFDocument59 pagesUPSC New Syllabus & Tips To Crack IAS Preliminary & Mains Exam PDFPrateek SahniNo ratings yet
- Kleiberit PUR 501 - FISPQDocument8 pagesKleiberit PUR 501 - FISPQYuri SouzaNo ratings yet
- Scarano 2009Document8 pagesScarano 2009Sabrina CaramNo ratings yet
- ESBIO 114 Module 8 BIOMESDocument17 pagesESBIO 114 Module 8 BIOMESMeah Angela GulacNo ratings yet
- Chapter 2 Envi SciDocument6 pagesChapter 2 Envi SciNadine PantiNo ratings yet
- ST 17 WindDocument10 pagesST 17 WindMarina ThomasNo ratings yet
- Temperate Grasslands OF South AmericaDocument41 pagesTemperate Grasslands OF South AmericaMario Tenorio MaldonadoNo ratings yet
- BiomesDocument26 pagesBiomesMariyam SiddiquiNo ratings yet
- Global Ecology: Chapter OutlineDocument25 pagesGlobal Ecology: Chapter OutlinehyNo ratings yet
- Review of Related LiteratureDocument12 pagesReview of Related LiteratureEllaNatividad0% (1)
- Enriched Advanced Ecology Notes 2020Document62 pagesEnriched Advanced Ecology Notes 2020aditi.ugandaNo ratings yet
- Asquith e Chang 2005 - DSDocument12 pagesAsquith e Chang 2005 - DSBeatrizNo ratings yet
- GGR 305 Week 1Document7 pagesGGR 305 Week 1ashNo ratings yet
- White Paper - CleanDocument10 pagesWhite Paper - Cleanapi-470802179No ratings yet
- Effects of Fire On Vegetation and Small Mammal Communities in A Mojave DesertDocument10 pagesEffects of Fire On Vegetation and Small Mammal Communities in A Mojave DesertRubens HayekNo ratings yet
- Tree Species Diversity, Composition and Aboveground Biomass Across Dry Forest Land-Cover Types in Coastal EcuadorDocument13 pagesTree Species Diversity, Composition and Aboveground Biomass Across Dry Forest Land-Cover Types in Coastal EcuadorLouis Paul KABELONGNo ratings yet
- Ecology CycleDocument30 pagesEcology CycleNiko Jay RojasNo ratings yet
- Asquith y Mejia - Chang-Mammals, Edge Effects, and The Loss of Tropical Forest Diversity-Ecology-2005Document13 pagesAsquith y Mejia - Chang-Mammals, Edge Effects, and The Loss of Tropical Forest Diversity-Ecology-2005braulioasensioNo ratings yet
- Site AnalysisDocument12 pagesSite AnalysisAntonny Christofer QlNo ratings yet
- Biogeographic Patterns and Conservation in The South American Cerrado: A Tropical Savanna HotspotDocument12 pagesBiogeographic Patterns and Conservation in The South American Cerrado: A Tropical Savanna HotspotANDRES RICARDO LOPEZ PRECIADONo ratings yet
- 1998 - The Global 200 Olson y DinersteinDocument15 pages1998 - The Global 200 Olson y DinersteinDALIANNA YICELL ARIZA CONCHILANo ratings yet
- Peopling Island Rainforests Global Trends From The Early Pleistocene To The Late HoloceneDocument26 pagesPeopling Island Rainforests Global Trends From The Early Pleistocene To The Late HoloceneRobby SunataNo ratings yet
- Hamer Et Al 2006 Carrion V FruitDocument9 pagesHamer Et Al 2006 Carrion V FruitKellyta RodriguezNo ratings yet
- Activity 3Document4 pagesActivity 3Slyther aceNo ratings yet
- Amazonia Is The Primary Source of Neotropical BiodiversityDocument6 pagesAmazonia Is The Primary Source of Neotropical BiodiversitySamuel FreitasNo ratings yet
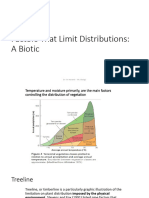
- Week - 5. Factors That Limit Distributions.a Biotic. NicheDocument31 pagesWeek - 5. Factors That Limit Distributions.a Biotic. NicheAnnisa VeterinerianNo ratings yet
- Genetic Effects of Rainforest Fragmentation in An Early Successional Tree (Elaeocarpus Grandis)Document9 pagesGenetic Effects of Rainforest Fragmentation in An Early Successional Tree (Elaeocarpus Grandis)secrettasteuhhNo ratings yet
- Guevara 2005Document8 pagesGuevara 2005Roger GuevaraNo ratings yet
- The Ecology and Evolution of Monito Del Monte (Fonturbel, 2022)Document17 pagesThe Ecology and Evolution of Monito Del Monte (Fonturbel, 2022)Gabriel Eduardo Mendoza RiquelmeNo ratings yet
- Riffle Algal and Macroinvertebrate Recovery From Wet-Seasonflood Disturbance in A Tropical Savannah RiverDocument18 pagesRiffle Algal and Macroinvertebrate Recovery From Wet-Seasonflood Disturbance in A Tropical Savannah RiverEverton Gustavo Miguel NevesNo ratings yet
- Blackwell Publishing Is Collaborating With JSTOR To Digitize, Preserve and Extend Access To Global Ecology and Biogeography LettersDocument22 pagesBlackwell Publishing Is Collaborating With JSTOR To Digitize, Preserve and Extend Access To Global Ecology and Biogeography LettersRanveer RajbharNo ratings yet
- Jordan 3e Chapter3 PDFDocument20 pagesJordan 3e Chapter3 PDFShane Franchezka GarciaNo ratings yet
- An Introduction To The Ecology of Desert SystemsDocument28 pagesAn Introduction To The Ecology of Desert SystemsInternational Journal of Innovative Science and Research TechnologyNo ratings yet
- Biological Conservation: Cameron D. Siler, Andrés Lira-Noriega, Rafe M. BrownDocument9 pagesBiological Conservation: Cameron D. Siler, Andrés Lira-Noriega, Rafe M. Brownmarie parfanNo ratings yet
- This Study Resource Was: Writing Portfolio Task 2 Paraphrase and Summary Form (Final)Document5 pagesThis Study Resource Was: Writing Portfolio Task 2 Paraphrase and Summary Form (Final)flz sfrNo ratings yet
- 17 MacroinvertebrateCommunities PDFDocument12 pages17 MacroinvertebrateCommunities PDFIJEAB JournalNo ratings yet
- Biodiversity and Healthy SocietyDocument12 pagesBiodiversity and Healthy SocietyKim CyrilNo ratings yet
- 121 1995 Biodiversity at Its UtmostDocument14 pages121 1995 Biodiversity at Its UtmostArghya PaulNo ratings yet
- Final Module EnviSciDocument55 pagesFinal Module EnviSciJeff Ray BarreraNo ratings yet
- 1 s2.0 S0367253021001171 MainDocument12 pages1 s2.0 S0367253021001171 MainValeria CasiqueNo ratings yet
- Amici & Nadkarni 2020 Differences in Epiphyte Biomass and CompositionDocument13 pagesAmici & Nadkarni 2020 Differences in Epiphyte Biomass and Compositionnayely.ggarciaNo ratings yet
- Definition CentralgrasslandDocument6 pagesDefinition Centralgrasslandapi-534620961No ratings yet
- Life Strategies of Mosses in Mt. Pulag, Benguet Province, PhilippinesDocument8 pagesLife Strategies of Mosses in Mt. Pulag, Benguet Province, PhilippinesRicky Justin NgoNo ratings yet
- Theacher GuideDocument4 pagesTheacher GuideAlejandro Lavid SaizNo ratings yet
- Macroalgae PDFDocument28 pagesMacroalgae PDFhanifazhar1811No ratings yet
- Best Animal BiomesDocument6 pagesBest Animal Biomessalman672003No ratings yet
- Diversity and Ecological Status of Bryophytes in Mt. Kitanglad, Bukidnon, PhilippinesDocument31 pagesDiversity and Ecological Status of Bryophytes in Mt. Kitanglad, Bukidnon, PhilippinesKennethRavanillaNo ratings yet
- Inger and Colwell 1977Document27 pagesInger and Colwell 19771803794dNo ratings yet
- Macrophytes: Ecology of Aquatic PlantsDocument18 pagesMacrophytes: Ecology of Aquatic PlantsQueng ElediaNo ratings yet
- Antonelli - Etal.2018.amazonina DiversityDocument6 pagesAntonelli - Etal.2018.amazonina Diversitycamila zapataNo ratings yet
- The Role of Macrophytes in Wetland EcosystemsDocument13 pagesThe Role of Macrophytes in Wetland EcosystemsEVELYN TABORDA MONCADANo ratings yet
- Journal of Ecology - 2003 - ImbertDocument10 pagesJournal of Ecology - 2003 - ImbertNicolas PeñarandaNo ratings yet
- The Association For Tropical Biology and ConservationDocument16 pagesThe Association For Tropical Biology and ConservationAlexis Centeno CuellarNo ratings yet
- Present State and Future of The Worlds Mangrove FDocument20 pagesPresent State and Future of The Worlds Mangrove F1658S.M. Atikul AlamNo ratings yet
- The Concept of Biomes - Interactive PresentationDocument70 pagesThe Concept of Biomes - Interactive PresentationjohnosborneNo ratings yet
- Biodiversity of Sri LankaDocument37 pagesBiodiversity of Sri LankaMonster AwakenedNo ratings yet
- Respuestas Post Incendio de Quercus HumboldtiiDocument12 pagesRespuestas Post Incendio de Quercus HumboldtiiJuan PerezNo ratings yet
- Teaching Activity Observing Victorias EcosystemsDocument3 pagesTeaching Activity Observing Victorias EcosystemsGeoff DammNo ratings yet
- Running Head: Maintaining Rainforest 1Document10 pagesRunning Head: Maintaining Rainforest 1HugsNo ratings yet
- Biodiversity: Explore the Diversity of Life on Earth with Environmental Science Activities for KidsFrom EverandBiodiversity: Explore the Diversity of Life on Earth with Environmental Science Activities for KidsNo ratings yet
- Synchronous Generators: Instructional ObjectivesDocument18 pagesSynchronous Generators: Instructional Objectivessanthosh2009No ratings yet
- Asma G.SDocument5 pagesAsma G.SAfia FaheemNo ratings yet
- 10 Science TP 11 1Document5 pages10 Science TP 11 1Ananaya BansalNo ratings yet
- Al55 66 Technical Manual v11 EngDocument60 pagesAl55 66 Technical Manual v11 EngProblem VelikiNo ratings yet
- Owner: PT. Baker Hughes User: Cok Gede Reza Description: Modified Safety Pin, Add. Handrail & Add. Anti Slip Step Stair at Pressure Test BayDocument3 pagesOwner: PT. Baker Hughes User: Cok Gede Reza Description: Modified Safety Pin, Add. Handrail & Add. Anti Slip Step Stair at Pressure Test BayMuhammad AlpianNo ratings yet
- Bata Shoe Company (Bangladesh) Ltd.Document6 pagesBata Shoe Company (Bangladesh) Ltd.Vurdalack666No ratings yet
- Green Building Toward Construction Sustainability: Energy Efficiency With Material and Design AspectsDocument11 pagesGreen Building Toward Construction Sustainability: Energy Efficiency With Material and Design AspectsghchgNo ratings yet
- Oil & Chemical Tanker Summer 7 - Prepurchase Survey Report - S&a 230098 PpsDocument62 pagesOil & Chemical Tanker Summer 7 - Prepurchase Survey Report - S&a 230098 Ppsp_k_sahuNo ratings yet
- Jayvee Dime,,,,project in Physics Light and SoundDocument25 pagesJayvee Dime,,,,project in Physics Light and SoundJayvee DimeNo ratings yet
- Data Sheet USB5 II 2019 05 ENDocument1 pageData Sheet USB5 II 2019 05 ENJanne LaineNo ratings yet
- Understanding Your Electricity Bill in PakistanDocument13 pagesUnderstanding Your Electricity Bill in PakistanGhayas Ud-din DarNo ratings yet
- Mouser TestsectionDocument33 pagesMouser TestsectionalltheloveintheworldNo ratings yet
- Metzeler TDB 2019 LRDocument246 pagesMetzeler TDB 2019 LRGabriel BruschiNo ratings yet
- Production of Biodegradable Plastics AsDocument6 pagesProduction of Biodegradable Plastics AsEbenezer EffisahNo ratings yet
- Bigbang PDFDocument772 pagesBigbang PDFLeanne Haddock100% (1)
- BW Inside CiscoDocument3 pagesBW Inside CiscoMunkhbayar BaadgaiNo ratings yet
- Cults and Temples of The Middle World (FG&G)Document77 pagesCults and Temples of The Middle World (FG&G)Paul SavvyNo ratings yet
- 9852 1880 01 Maintenance Instructions ROC L7CR IIDocument102 pages9852 1880 01 Maintenance Instructions ROC L7CR IIphongNo ratings yet
- Geometry - Olympiad Material BY DR SHYAM SUNDAR AGRAWALDocument67 pagesGeometry - Olympiad Material BY DR SHYAM SUNDAR AGRAWALdrssagrawalNo ratings yet
- Macro Chapter 7Document12 pagesMacro Chapter 7Mary Jane PelaezNo ratings yet
- Coal Ash Dewatering SolutionDocument23 pagesCoal Ash Dewatering Solution류태하No ratings yet
- LECLANCHE Cell Datasheet GL60 GNMC 60ahDocument2 pagesLECLANCHE Cell Datasheet GL60 GNMC 60ahChaitanyaNo ratings yet
- BEL PE Question Papers For ECE With Answers - 6 Knowledge AddaDocument14 pagesBEL PE Question Papers For ECE With Answers - 6 Knowledge AddaVishwanand ThombareNo ratings yet
- 4 Poisonous & Venomous AnimalsDocument47 pages4 Poisonous & Venomous AnimalsAnyi Yulieth AMPUDIA MURILLONo ratings yet
- Electronics Cooling: Mechanical Power Engineering DeptDocument22 pagesElectronics Cooling: Mechanical Power Engineering DeptneilNo ratings yet
- Hillfoot Data Sheets V5 1Document4 pagesHillfoot Data Sheets V5 1Kristijan IlicNo ratings yet
- Combustion Engineering, Heat Transfer, Refrigeration Engineering, & Air ConditioningDocument17 pagesCombustion Engineering, Heat Transfer, Refrigeration Engineering, & Air ConditioningNicole Mae AllosadaNo ratings yet