Download as pdf or txt
You might also like
- Science Week Proposal 2 JuneDocument26 pagesScience Week Proposal 2 Junecsanjeevan100% (1)
- Architects Illustrated Pocket DictionaryDocument558 pagesArchitects Illustrated Pocket DictionaryFlavia ValNo ratings yet
- Catecholamine: Epinephrine (Adrenaline)Document7 pagesCatecholamine: Epinephrine (Adrenaline)Unggul YudhaNo ratings yet
- Amine NeurotransmittersDocument7 pagesAmine NeurotransmittersFrancisco BecerraNo ratings yet
- NP3 PDFDocument30 pagesNP3 PDFRajat AgrawalNo ratings yet
- Central Nervous System StimulantsDocument10 pagesCentral Nervous System StimulantsCristian F AponteNo ratings yet
- Faktor Neurobiologi: Qara Syifa Fachrani 1610211039Document22 pagesFaktor Neurobiologi: Qara Syifa Fachrani 1610211039Qara Syifa FNo ratings yet
- Adrenergic Agents SeminarDocument48 pagesAdrenergic Agents SeminarTheBoss 20No ratings yet
- 5-Hydroxytryptophan: Michael T. Murray, NDDocument14 pages5-Hydroxytryptophan: Michael T. Murray, NDDoina DoniciNo ratings yet
- MedchemIII PHSC5100 2018 .PostDocument43 pagesMedchemIII PHSC5100 2018 .PostMamta ShindeNo ratings yet
- Toxicity of Amphetamines: An UpdateDocument65 pagesToxicity of Amphetamines: An UpdateMariana MadiaNo ratings yet
- roledopamineOMCL2016 9730467Document13 pagesroledopamineOMCL2016 9730467gbautista.fuerteNo ratings yet
- CH 10Document26 pagesCH 10osama2010bNo ratings yet
- Biosynthetic Pathways For Catecholamines and in The: Trace Amines Human BrainDocument3 pagesBiosynthetic Pathways For Catecholamines and in The: Trace Amines Human BrainVincent Paul SantosNo ratings yet
- 3 ANSDrugsDocument75 pages3 ANSDrugsLinearNo ratings yet
- Tugas Michelle 2Document7 pagesTugas Michelle 2andre nicholasNo ratings yet
- Adrenergic DrugsDocument21 pagesAdrenergic DrugsArturo RinconNo ratings yet
- Unit-2 Medicinal Chemistry 4th Sem B.pharma Module Bright MedicoDocument26 pagesUnit-2 Medicinal Chemistry 4th Sem B.pharma Module Bright Medicoparmarkeval1610No ratings yet
- Bylund 2014Document3 pagesBylund 2014Javier RiveraNo ratings yet
- Substituted Phenethylamine - WikipediaDocument4 pagesSubstituted Phenethylamine - WikipediaSuzane RosaNo ratings yet
- Chapter 1 Adrenergic Drugs FinalDocument52 pagesChapter 1 Adrenergic Drugs FinalShivKumarNo ratings yet
- Reviewer of A PsychologyDocument55 pagesReviewer of A PsychologyCAMMILLE EDZ FERRAS SELONGNo ratings yet
- Cpy in The Metabolism of DrugsDocument2 pagesCpy in The Metabolism of DrugsAngie CeronNo ratings yet
- Adrenaline PresentationDocument12 pagesAdrenaline PresentationAshley MudzingwaNo ratings yet
- Biochemistry of NeurotransmittersDocument7 pagesBiochemistry of NeurotransmittersHyphophysis 2015No ratings yet
- Adrenergic and Anti-Adrenergic Drugs: Manju PrathapDocument43 pagesAdrenergic and Anti-Adrenergic Drugs: Manju PrathapManju PrathapNo ratings yet
- Adrenal GlandDocument49 pagesAdrenal GlandRaphael AnajeNo ratings yet
- Week 16 - Adrenergic AgentsDocument33 pagesWeek 16 - Adrenergic AgentsCham MontimanNo ratings yet
- Adrenergic 1Document56 pagesAdrenergic 1Elaf SherzadNo ratings yet
- Biosynthesis of Alkaloid Natural Products 5.1. Alkaloids Are Derived From Amino AcidsDocument14 pagesBiosynthesis of Alkaloid Natural Products 5.1. Alkaloids Are Derived From Amino Acidsharishkumar kakrani100% (2)
- Antipsychotics 1Document25 pagesAntipsychotics 1bcloverasta905No ratings yet
- Adrenergik-Antiadrenergik 2013Document63 pagesAdrenergik-Antiadrenergik 2013MeldianaNo ratings yet
- MSB 202: Neurolocomotor: Neurotransmitters - Overview of Metabolism (Anabolism and Catabolism)Document42 pagesMSB 202: Neurolocomotor: Neurotransmitters - Overview of Metabolism (Anabolism and Catabolism)Jacob MasikaNo ratings yet
- J. Biol. Chem.-1958-Axelrod-702-5Document5 pagesJ. Biol. Chem.-1958-Axelrod-702-5Elvira Claveluz Aparicio AquiseNo ratings yet
- GFDGFDGFDGDocument11 pagesGFDGFDGFDGلو ترىNo ratings yet
- Yagiela John A Et Al Pharmacology and Therapeutics For Denti-3Document1 pageYagiela John A Et Al Pharmacology and Therapeutics For Denti-3AntonPurpurovNo ratings yet
- NeurotransmittersDocument21 pagesNeurotransmittersYannie GomezNo ratings yet
- Catecholamines: (Dopamine (DA), Norepinephrine (NE), Epinephrine (EPI) )Document30 pagesCatecholamines: (Dopamine (DA), Norepinephrine (NE), Epinephrine (EPI) )Imrana AamirNo ratings yet
- Physiology, Catecholamines: Stephen Paravati Alan Rosani Steven J. WarringtonDocument2 pagesPhysiology, Catecholamines: Stephen Paravati Alan Rosani Steven J. WarringtonVincent Paul SantosNo ratings yet
- NEPHAR 305 Antiepileptic - 10Document17 pagesNEPHAR 305 Antiepileptic - 10Raju NiraulaNo ratings yet
- 220 Alkaloids 4 Lecture 12Document27 pages220 Alkaloids 4 Lecture 12حسن عباس حسن عباس الحسونNo ratings yet
- Serotonin (5-Hydroxytryptamine)Document74 pagesSerotonin (5-Hydroxytryptamine)matchees-gone rogueNo ratings yet
- Pharmacol Ogy PHL 313: Chapter 5-6-7 Drugs Affecting The Autonomic Nervous System (ANS)Document14 pagesPharmacol Ogy PHL 313: Chapter 5-6-7 Drugs Affecting The Autonomic Nervous System (ANS)Abdullah Al-TuraifiNo ratings yet
- Hormones 1 2015 BDocument62 pagesHormones 1 2015 BJonathan JohnsonNo ratings yet
- Autonomic Nervous SystemDocument54 pagesAutonomic Nervous SystemEzio SartoraNo ratings yet
- Synthesis and Significance of 5-Hydroxy Tryptamine (Or) SerotoninDocument10 pagesSynthesis and Significance of 5-Hydroxy Tryptamine (Or) SerotoninVikas SharmaNo ratings yet
- L9 Sympathomimetics Lytics REVISED 2017 PDFDocument49 pagesL9 Sympathomimetics Lytics REVISED 2017 PDFVea AngelesNo ratings yet
- Dopamine, Serotonine, Epinephrine, and NorephinephrineDocument28 pagesDopamine, Serotonine, Epinephrine, and Norephinephrinehafshah100% (1)
- ANFETAMINASDocument10 pagesANFETAMINASIza ÁlvarezNo ratings yet
- Extracted Pages From Basic - and - Clinical - Pharmacology - Fourteen - 2Document8 pagesExtracted Pages From Basic - and - Clinical - Pharmacology - Fourteen - 2ed trsgf rgsfNo ratings yet
- Introduction To Nervous System - NeurotransmittersDocument19 pagesIntroduction To Nervous System - NeurotransmittersKate TaylorNo ratings yet
- Copmpounds Formed From Tyrosine and PhenylalanineDocument21 pagesCopmpounds Formed From Tyrosine and PhenylalaninesandigawadnehaNo ratings yet
- Metabolisme Xenobiotik Metabolisme Xenobiotik: Toxicants BiotrasformationDocument42 pagesMetabolisme Xenobiotik Metabolisme Xenobiotik: Toxicants BiotrasformationTRIA SAPUTRA100% (1)
- Nucleotide Biosynthesis Lecture 2017sDocument23 pagesNucleotide Biosynthesis Lecture 2017sSaad KazmiNo ratings yet
- Adrenergic Drugs 10 - 2007Document57 pagesAdrenergic Drugs 10 - 2007أمجد محمدNo ratings yet
- NorepinephrineDocument9 pagesNorepinephrineDimitris KaragkiozidisNo ratings yet
- Extracted Pages From Basic - and - Clinical - Pharmacology - Fourteen - 2Document11 pagesExtracted Pages From Basic - and - Clinical - Pharmacology - Fourteen - 2ed trsgf rgsfNo ratings yet
- CC2 - Therapeutic Drugs - Drugs of Abuse&PoisonsNEWDocument34 pagesCC2 - Therapeutic Drugs - Drugs of Abuse&PoisonsNEWYlia MastarsNo ratings yet
- 400 3 NarcoticsDocument27 pages400 3 Narcoticsfirdaus92No ratings yet
- Neurotransmitters - Overview of Synthesis and MetabolismDocument31 pagesNeurotransmitters - Overview of Synthesis and MetabolismnehaNo ratings yet
- Information Kucat CBT 2019 Sos Soe1562662484Document8 pagesInformation Kucat CBT 2019 Sos Soe1562662484nabin hamalNo ratings yet
- Kathmandu Medical College: Application For The Mbbs Programme 201 - 201Document3 pagesKathmandu Medical College: Application For The Mbbs Programme 201 - 201nabin hamalNo ratings yet
- Nepal Health Research CouncilDocument15 pagesNepal Health Research Councilnabin hamalNo ratings yet
- Enlarged Prostate Booklet PDFDocument52 pagesEnlarged Prostate Booklet PDFnabin hamalNo ratings yet
- JPT PDFDocument11 pagesJPT PDFnabin hamalNo ratings yet
- Engine Valve DesignsDocument6 pagesEngine Valve DesignsGhanvir SinghNo ratings yet
- Cherry 1900 Bulbing Blind Rivet: SPS Fastener Division, ADocument4 pagesCherry 1900 Bulbing Blind Rivet: SPS Fastener Division, AДенис СергеевичNo ratings yet
- An Outline of The Life and Works of Col Paul Revere (1901)Document164 pagesAn Outline of The Life and Works of Col Paul Revere (1901)liketoreadNo ratings yet
- Module 8 Neo BehaviorismDocument11 pagesModule 8 Neo BehaviorismDenyseNo ratings yet
- Ice Cream Frozen Dessrt PDFDocument145 pagesIce Cream Frozen Dessrt PDFSreelekshmi LechuNo ratings yet
- Ajay Kumar Shukla - CVDocument6 pagesAjay Kumar Shukla - CVdkannanapkNo ratings yet
- Fairies, Dragons & Witchblood - Rade RisticDocument4 pagesFairies, Dragons & Witchblood - Rade RisticChristopher A Ashbrook100% (1)
- Hyper-V Virtual Machine Connection - Microsoft DocsDocument5 pagesHyper-V Virtual Machine Connection - Microsoft DocsDeepak KumarNo ratings yet
- FactSheet DecathlonDocument2 pagesFactSheet DecathlonAli RamosNo ratings yet
- Endoscopy Patient Post Care FAQsDocument1 pageEndoscopy Patient Post Care FAQsMarie-Andree Roy-SajjaNo ratings yet
- Ratio Detector: SNSCT 16Ec304-Communication Theory Unit-3 K.Sumathi, Ap/EceDocument2 pagesRatio Detector: SNSCT 16Ec304-Communication Theory Unit-3 K.Sumathi, Ap/EceChandu NanduNo ratings yet
- Thesis CoutureDocument6 pagesThesis Couturemariapolitepalmdale100% (2)
- Brosur Buffet Package TelView 2013Document8 pagesBrosur Buffet Package TelView 2013gearkitNo ratings yet
- Final-Past Exam QuestionsDocument8 pagesFinal-Past Exam QuestionsKhánh Linh Nguyễn NgọcNo ratings yet
- Janma Vighatika GrahaDocument5 pagesJanma Vighatika Grahatimsmith1081574No ratings yet
- Isotrend Ir 192 Brochure PDFDocument2 pagesIsotrend Ir 192 Brochure PDFSafiq UddinNo ratings yet
- Jaguar XK - TYPE 2003 Model Year Electrical GuideDocument69 pagesJaguar XK - TYPE 2003 Model Year Electrical GuideCesar ArteagaNo ratings yet
- 12 English - Writing Skills-AdvertisementsDocument13 pages12 English - Writing Skills-AdvertisementsSiddhant KapoorNo ratings yet
- Stress Analysis of Steam Piping SystemDocument6 pagesStress Analysis of Steam Piping SystemDaniel Camilo SalamancaNo ratings yet
- Cells of Immune System Notes 2Document70 pagesCells of Immune System Notes 2Sudeeksha Ravikoti100% (1)
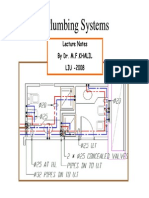
- Plumbing 2008 Includes FU Calculation Fire Fighting..Document203 pagesPlumbing 2008 Includes FU Calculation Fire Fighting..michelghanem100% (2)
- Alternating Current and VoltageDocument5 pagesAlternating Current and VoltageRohit SahuNo ratings yet
- Цацулин CQC Mag 2001-06 J - eng PDFDocument1 pageЦацулин CQC Mag 2001-06 J - eng PDFBudo MediaNo ratings yet
- Department of Technical EducationDocument24 pagesDepartment of Technical EducationGagan SNo ratings yet
- 01 Masonry Grout Filler - Group 180821Document2 pages01 Masonry Grout Filler - Group 180821Maha MuflehNo ratings yet
- TPC-1071H - 1271H - 1571H - 1771H - User Manual - Ed3Document88 pagesTPC-1071H - 1271H - 1571H - 1771H - User Manual - Ed3DeniMestiWidiantoNo ratings yet
- YS VIII Steam ArtbookDocument19 pagesYS VIII Steam ArtbookTuukka MäkinenNo ratings yet