
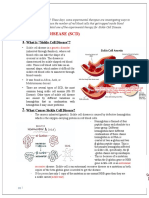
Hemoglobinopathies (Hemoglobin Disorders)
Hemoglobinopathies (Hemoglobin Disorders)
You might also like
- The Uninformed Coach RevisedDocument11 pagesThe Uninformed Coach Revisedapi-292411774No ratings yet
- Biology Investigatory PROJECT ThallassemiaDocument35 pagesBiology Investigatory PROJECT ThallassemiaBiswajit Rout85% (13)
- Assignment On ThalassemiaDocument5 pagesAssignment On ThalassemiaMuniraNo ratings yet
- HemoglobinDocument5 pagesHemoglobinAyioKunNo ratings yet
- HemoglobinopathiesDocument37 pagesHemoglobinopathiesBabak Nami100% (1)
- Sickle Cell Disease in PregnancyDocument18 pagesSickle Cell Disease in Pregnancyapi-370504667% (3)
- Hemoglobin SynthesisDocument4 pagesHemoglobin Synthesismadison DeliNo ratings yet
- 02 HemoglobinDocument78 pages02 HemoglobinpixiedustNo ratings yet
- HemoglobinDocument41 pagesHemoglobinSri Ram 07No ratings yet
- Hemoglobinopatzhy PDFDocument16 pagesHemoglobinopatzhy PDFJoy CuanoNo ratings yet
- Introduction To Hemoglobinopathies Diagnostics On HPLC by P.C. GiordanoDocument4 pagesIntroduction To Hemoglobinopathies Diagnostics On HPLC by P.C. GiordanoUMMID WashimNo ratings yet
- Hemoglobinopathies & Thalasemia Learning ObjectivesDocument10 pagesHemoglobinopathies & Thalasemia Learning ObjectivesFatima AhmedNo ratings yet
- Thalassemia: Submitted By: Jovan Pierre C. Ouano Submitted To: Mark Gil T. DacutanDocument8 pagesThalassemia: Submitted By: Jovan Pierre C. Ouano Submitted To: Mark Gil T. DacutanJvnpierre AberricanNo ratings yet
- 7 HemoglobinopathiesDocument34 pages7 HemoglobinopathiesAhmad SobihNo ratings yet
- Thalassaemia: Definition, Classification, Pathogenesis and Complications of This DisorderDocument37 pagesThalassaemia: Definition, Classification, Pathogenesis and Complications of This DisorderSheila MichaelsNo ratings yet
- Hemoglobin ElectrophoresisDocument22 pagesHemoglobin ElectrophoresisMohamed IbrahimNo ratings yet
- Background: Alpha ThalassemiaDocument24 pagesBackground: Alpha ThalassemiacristieristiieNo ratings yet
- Hem AssignmentDocument5 pagesHem Assignmentmalar_km43No ratings yet
- Haemoglobinopathies - ThalassaemiaDocument49 pagesHaemoglobinopathies - ThalassaemiaRaymond LundaNo ratings yet
- Pages From Download-4Document1 pagePages From Download-4Ra,i Al HaddadNo ratings yet
- PB 0783Document9 pagesPB 0783Mona HelouNo ratings yet
- 4 Genetics DiseaseDocument40 pages4 Genetics Diseasemouid0.2003No ratings yet
- Thalassemia and Related HemoglobinopathiesDocument6 pagesThalassemia and Related HemoglobinopathiesMelly SyafridaNo ratings yet
- Hemoglobinopathy - WikipediaDocument9 pagesHemoglobinopathy - WikipediaEslam NassarNo ratings yet
- Beta ThalassemiaDocument17 pagesBeta ThalassemiaNorman DamaaNo ratings yet
- THALASSEMIADocument33 pagesTHALASSEMIAMirzi Cuison100% (1)
- ThalassemiaDocument3 pagesThalassemiaArmandoNo ratings yet
- Hemoglobine SynthesisDocument4 pagesHemoglobine SynthesisNOVRILIANo ratings yet
- Biochemistry - Hemoglbin SynthesisDocument4 pagesBiochemistry - Hemoglbin SynthesisBildad Felix KirandeNo ratings yet
- Alpha and Beta ThalassemiaDocument6 pagesAlpha and Beta ThalassemiaSi vis pacem...100% (1)
- Hemoglobinopathies and Thalassemias Lin OswariDocument16 pagesHemoglobinopathies and Thalassemias Lin OswariNOVRILIANo ratings yet
- Lecture 2Document35 pagesLecture 2Adams ZarawuNo ratings yet
- What Is Sickle Cell DiseaseDocument11 pagesWhat Is Sickle Cell DiseasePhương Ly LêNo ratings yet
- ThalassemiaDocument5 pagesThalassemiaVeronica Wong Huey ShinNo ratings yet
- HB Bart Fact SheetDocument2 pagesHB Bart Fact Sheetstrawberry pieNo ratings yet
- Hemoglobin Op A Thy Is A Kind of Genetic Defect That Results in Abnormal Structure of One of The Globin Chains of The Hemoglobin MoleculeDocument3 pagesHemoglobin Op A Thy Is A Kind of Genetic Defect That Results in Abnormal Structure of One of The Globin Chains of The Hemoglobin MoleculeIkram ShinwariNo ratings yet
- Hemoglobinopathies and Carbon Monoxide PoisoningDocument45 pagesHemoglobinopathies and Carbon Monoxide PoisoningBea SamonteNo ratings yet
- Genetic Disorders of Haeoglobin - Haemoglobinopathies - 200622Document22 pagesGenetic Disorders of Haeoglobin - Haemoglobinopathies - 200622yussufNo ratings yet
- HemoglobinopathiesDocument26 pagesHemoglobinopathiesSanjey GaneshNo ratings yet
- Thalassemias: Alissa Martin,, Alexis A. ThompsonDocument9 pagesThalassemias: Alissa Martin,, Alexis A. ThompsonAmada RosalesNo ratings yet
- Sickle Cell Anemia and Thalassemias: Paul R. EarlDocument21 pagesSickle Cell Anemia and Thalassemias: Paul R. EarlNavisatul MutmainahNo ratings yet
- Thalassemia: by DR PP GevaoDocument46 pagesThalassemia: by DR PP GevaoAbubakar JallohNo ratings yet
- Erythrocyte Disorders in The Perinatal Period in AdversePregnancy Outcome and The Fetus:NeonateDocument12 pagesErythrocyte Disorders in The Perinatal Period in AdversePregnancy Outcome and The Fetus:Neonatethanhhien900No ratings yet
- Hematology I - Hemoglobin: Physiologic FormsDocument4 pagesHematology I - Hemoglobin: Physiologic FormsAldren BeliberNo ratings yet
- Hemoglobinopathies and ThalassemiasDocument10 pagesHemoglobinopathies and ThalassemiashartNo ratings yet
- Detection and Quantitation of Normal and Variant Haemoglobins: An Analytical ReviewDocument15 pagesDetection and Quantitation of Normal and Variant Haemoglobins: An Analytical ReviewAmanda MatthewsNo ratings yet
- Introduction To HemoglobinopathiesDocument12 pagesIntroduction To HemoglobinopathiesMichael MazvidzaNo ratings yet
- Alpha and Betha Thalassemia (Muncie, 2009)Document8 pagesAlpha and Betha Thalassemia (Muncie, 2009)widyafandriNo ratings yet
- Thalassemiafinal 111212142013 Phpapp02 130321172427 Phpapp01Document35 pagesThalassemiafinal 111212142013 Phpapp02 130321172427 Phpapp01MUHAMMAD WAQAS TARIQNo ratings yet
- ThalassaemiasDocument8 pagesThalassaemiasSalim UddinkhanNo ratings yet
- Hemoglobin and Iron Metabolism: Transport of Respiratory GasesDocument6 pagesHemoglobin and Iron Metabolism: Transport of Respiratory GasesAnusuya SNo ratings yet
- ThalassaemiasDocument8 pagesThalassaemiasSalim UddinkhanNo ratings yet
- Thalasemia MedscapeDocument21 pagesThalasemia MedscapeFenty IswarNo ratings yet
- THALASSEMIADocument30 pagesTHALASSEMIAjismi vallachiraNo ratings yet
- Hematology Lectures 1 5 DR - TuyDocument10 pagesHematology Lectures 1 5 DR - TuyMiguel Cuevas DolotNo ratings yet
- Thalassaemias PDFDocument3 pagesThalassaemias PDFNidia100% (1)
- Hemoglobinopathies Occupy A Special Place in Human Genetics For Many ReasonsDocument17 pagesHemoglobinopathies Occupy A Special Place in Human Genetics For Many ReasonsjipmerbiochemNo ratings yet
- Pages From Download-2Document1 pagePages From Download-2Ra,i Al HaddadNo ratings yet
- Heme ChemistryDocument23 pagesHeme ChemistryShereen KaddafyNo ratings yet
- Hemoglobin Opa ThiesDocument34 pagesHemoglobin Opa ThiesFebri fitraNo ratings yet
- HemoglobinDocument7 pagesHemoglobinNinik Triayu S100% (1)
- A Guide To Life!: - and List To Live byDocument6 pagesA Guide To Life!: - and List To Live byBravan AliennNo ratings yet
- Computer Data Storage-1Document9 pagesComputer Data Storage-1Bravan AliennNo ratings yet
- 6 ThrombosisDocument27 pages6 ThrombosisBravan AliennNo ratings yet
- Vascularized Tissue To Injurious Agent A Protective ResponseDocument13 pagesVascularized Tissue To Injurious Agent A Protective ResponseBravan AliennNo ratings yet
- Cellular Events: A Critical Function of InflammationDocument21 pagesCellular Events: A Critical Function of InflammationBravan AliennNo ratings yet
- Bible Truth and The TruthDocument228 pagesBible Truth and The TruthJeff Stutz100% (1)
- Sickle Cell AnemiaDocument23 pagesSickle Cell AnemiaJesmin_36No ratings yet
- ThlasmiaDocument7 pagesThlasmiaSonia AhmedNo ratings yet
- ThalcemiaDocument33 pagesThalcemiaRahul Kumar DiwakarNo ratings yet
- Blood Typing ExperimentDocument6 pagesBlood Typing ExperimentAdib SadmanNo ratings yet
- Study On Hemoglobinopathies and G6PD Deficiency in Terai Districts of NepalDocument30 pagesStudy On Hemoglobinopathies and G6PD Deficiency in Terai Districts of NepalSaroj BhattaraiNo ratings yet
- Note That There Are Several Different "Basic Blast" Programs Available at Ncbi (Including Nucleotide Blast, Protein Blast, and Blastx)Document10 pagesNote That There Are Several Different "Basic Blast" Programs Available at Ncbi (Including Nucleotide Blast, Protein Blast, and Blastx)Raj Kumar SoniNo ratings yet
- Shivam Biology ProjectDocument18 pagesShivam Biology ProjectPankaj SolankiNo ratings yet
- Ann Druyan Talks About Carl Sagan and Science, Religion, Wonder, Awe - . .Document23 pagesAnn Druyan Talks About Carl Sagan and Science, Religion, Wonder, Awe - . .stephenking0078916No ratings yet
- AnemiaDocument33 pagesAnemiaJasmine Thabith100% (5)
- Case StudyDocument42 pagesCase StudyChristine Joy Ilao PasnoNo ratings yet
- Hereditary AnemiaDocument77 pagesHereditary AnemiaAravindh SivaNo ratings yet
- THALASSEMIADocument49 pagesTHALASSEMIAMunish DograNo ratings yet
- Army Public School Gopalpur: Chapter-5 Principles of Inheritance and Variation NotesDocument7 pagesArmy Public School Gopalpur: Chapter-5 Principles of Inheritance and Variation NotesAshok KumarNo ratings yet
- KVS Study Material Biology Class 12Document189 pagesKVS Study Material Biology Class 12Akshatha Nayak60% (5)
- Chapter 6 Hema 1Document7 pagesChapter 6 Hema 1EMAN SALEHNo ratings yet
- Sickle Cell AnemiaDocument37 pagesSickle Cell Anemiahazelposis75% (4)
- 3 ThalassemiaDocument20 pages3 ThalassemiaMARYLINENo ratings yet
- Dr. Silvanus Chakra PuspitaDocument74 pagesDr. Silvanus Chakra PuspitaDewi PratiwiNo ratings yet
- Structure, Types and Various Methods For Estimation of HaemoglobinDocument49 pagesStructure, Types and Various Methods For Estimation of HaemoglobinChandra ShekharNo ratings yet
- 12 Biology Revision Questions 2017 18 Chapter 5 PDFDocument15 pages12 Biology Revision Questions 2017 18 Chapter 5 PDFFaiz Alam100% (1)
- Bioinformatics-II (BIF501) : Assignment No.01Document15 pagesBioinformatics-II (BIF501) : Assignment No.01MehboobNo ratings yet
- DrepanocitozaDocument11 pagesDrepanocitozaElisa TelharajNo ratings yet
- AnemiaDocument145 pagesAnemiaTegegne WorkNo ratings yet
- Bioinformatic Analysis of HBBDocument1 pageBioinformatic Analysis of HBBfrank2689No ratings yet
- Background: Alpha ThalassemiaDocument24 pagesBackground: Alpha ThalassemiacristieristiieNo ratings yet
- 2 - Diagnosticul Molecular - Hype or HopeDocument57 pages2 - Diagnosticul Molecular - Hype or HopeSorin LazarNo ratings yet


























































































Download as doc, pdf, or txt
You might also like
- The Uninformed Coach RevisedDocument11 pagesThe Uninformed Coach Revisedapi-292411774No ratings yet
- Biology Investigatory PROJECT ThallassemiaDocument35 pagesBiology Investigatory PROJECT ThallassemiaBiswajit Rout85% (13)
- Assignment On ThalassemiaDocument5 pagesAssignment On ThalassemiaMuniraNo ratings yet
- HemoglobinDocument5 pagesHemoglobinAyioKunNo ratings yet
- HemoglobinopathiesDocument37 pagesHemoglobinopathiesBabak Nami100% (1)
- Sickle Cell Disease in PregnancyDocument18 pagesSickle Cell Disease in Pregnancyapi-370504667% (3)
- Hemoglobin SynthesisDocument4 pagesHemoglobin Synthesismadison DeliNo ratings yet
- 02 HemoglobinDocument78 pages02 HemoglobinpixiedustNo ratings yet
- HemoglobinDocument41 pagesHemoglobinSri Ram 07No ratings yet
- Hemoglobinopatzhy PDFDocument16 pagesHemoglobinopatzhy PDFJoy CuanoNo ratings yet
- Introduction To Hemoglobinopathies Diagnostics On HPLC by P.C. GiordanoDocument4 pagesIntroduction To Hemoglobinopathies Diagnostics On HPLC by P.C. GiordanoUMMID WashimNo ratings yet
- Hemoglobinopathies & Thalasemia Learning ObjectivesDocument10 pagesHemoglobinopathies & Thalasemia Learning ObjectivesFatima AhmedNo ratings yet
- Thalassemia: Submitted By: Jovan Pierre C. Ouano Submitted To: Mark Gil T. DacutanDocument8 pagesThalassemia: Submitted By: Jovan Pierre C. Ouano Submitted To: Mark Gil T. DacutanJvnpierre AberricanNo ratings yet
- 7 HemoglobinopathiesDocument34 pages7 HemoglobinopathiesAhmad SobihNo ratings yet
- Thalassaemia: Definition, Classification, Pathogenesis and Complications of This DisorderDocument37 pagesThalassaemia: Definition, Classification, Pathogenesis and Complications of This DisorderSheila MichaelsNo ratings yet
- Hemoglobin ElectrophoresisDocument22 pagesHemoglobin ElectrophoresisMohamed IbrahimNo ratings yet
- Background: Alpha ThalassemiaDocument24 pagesBackground: Alpha ThalassemiacristieristiieNo ratings yet
- Hem AssignmentDocument5 pagesHem Assignmentmalar_km43No ratings yet
- Haemoglobinopathies - ThalassaemiaDocument49 pagesHaemoglobinopathies - ThalassaemiaRaymond LundaNo ratings yet
- Pages From Download-4Document1 pagePages From Download-4Ra,i Al HaddadNo ratings yet
- PB 0783Document9 pagesPB 0783Mona HelouNo ratings yet
- 4 Genetics DiseaseDocument40 pages4 Genetics Diseasemouid0.2003No ratings yet
- Thalassemia and Related HemoglobinopathiesDocument6 pagesThalassemia and Related HemoglobinopathiesMelly SyafridaNo ratings yet
- Hemoglobinopathy - WikipediaDocument9 pagesHemoglobinopathy - WikipediaEslam NassarNo ratings yet
- Beta ThalassemiaDocument17 pagesBeta ThalassemiaNorman DamaaNo ratings yet
- THALASSEMIADocument33 pagesTHALASSEMIAMirzi Cuison100% (1)
- ThalassemiaDocument3 pagesThalassemiaArmandoNo ratings yet
- Hemoglobine SynthesisDocument4 pagesHemoglobine SynthesisNOVRILIANo ratings yet
- Biochemistry - Hemoglbin SynthesisDocument4 pagesBiochemistry - Hemoglbin SynthesisBildad Felix KirandeNo ratings yet
- Alpha and Beta ThalassemiaDocument6 pagesAlpha and Beta ThalassemiaSi vis pacem...100% (1)
- Hemoglobinopathies and Thalassemias Lin OswariDocument16 pagesHemoglobinopathies and Thalassemias Lin OswariNOVRILIANo ratings yet
- Lecture 2Document35 pagesLecture 2Adams ZarawuNo ratings yet
- What Is Sickle Cell DiseaseDocument11 pagesWhat Is Sickle Cell DiseasePhương Ly LêNo ratings yet
- ThalassemiaDocument5 pagesThalassemiaVeronica Wong Huey ShinNo ratings yet
- HB Bart Fact SheetDocument2 pagesHB Bart Fact Sheetstrawberry pieNo ratings yet
- Hemoglobin Op A Thy Is A Kind of Genetic Defect That Results in Abnormal Structure of One of The Globin Chains of The Hemoglobin MoleculeDocument3 pagesHemoglobin Op A Thy Is A Kind of Genetic Defect That Results in Abnormal Structure of One of The Globin Chains of The Hemoglobin MoleculeIkram ShinwariNo ratings yet
- Hemoglobinopathies and Carbon Monoxide PoisoningDocument45 pagesHemoglobinopathies and Carbon Monoxide PoisoningBea SamonteNo ratings yet
- Genetic Disorders of Haeoglobin - Haemoglobinopathies - 200622Document22 pagesGenetic Disorders of Haeoglobin - Haemoglobinopathies - 200622yussufNo ratings yet
- HemoglobinopathiesDocument26 pagesHemoglobinopathiesSanjey GaneshNo ratings yet
- Thalassemias: Alissa Martin,, Alexis A. ThompsonDocument9 pagesThalassemias: Alissa Martin,, Alexis A. ThompsonAmada RosalesNo ratings yet
- Sickle Cell Anemia and Thalassemias: Paul R. EarlDocument21 pagesSickle Cell Anemia and Thalassemias: Paul R. EarlNavisatul MutmainahNo ratings yet
- Thalassemia: by DR PP GevaoDocument46 pagesThalassemia: by DR PP GevaoAbubakar JallohNo ratings yet
- Erythrocyte Disorders in The Perinatal Period in AdversePregnancy Outcome and The Fetus:NeonateDocument12 pagesErythrocyte Disorders in The Perinatal Period in AdversePregnancy Outcome and The Fetus:Neonatethanhhien900No ratings yet
- Hematology I - Hemoglobin: Physiologic FormsDocument4 pagesHematology I - Hemoglobin: Physiologic FormsAldren BeliberNo ratings yet
- Hemoglobinopathies and ThalassemiasDocument10 pagesHemoglobinopathies and ThalassemiashartNo ratings yet
- Detection and Quantitation of Normal and Variant Haemoglobins: An Analytical ReviewDocument15 pagesDetection and Quantitation of Normal and Variant Haemoglobins: An Analytical ReviewAmanda MatthewsNo ratings yet
- Introduction To HemoglobinopathiesDocument12 pagesIntroduction To HemoglobinopathiesMichael MazvidzaNo ratings yet
- Alpha and Betha Thalassemia (Muncie, 2009)Document8 pagesAlpha and Betha Thalassemia (Muncie, 2009)widyafandriNo ratings yet
- Thalassemiafinal 111212142013 Phpapp02 130321172427 Phpapp01Document35 pagesThalassemiafinal 111212142013 Phpapp02 130321172427 Phpapp01MUHAMMAD WAQAS TARIQNo ratings yet
- ThalassaemiasDocument8 pagesThalassaemiasSalim UddinkhanNo ratings yet
- Hemoglobin and Iron Metabolism: Transport of Respiratory GasesDocument6 pagesHemoglobin and Iron Metabolism: Transport of Respiratory GasesAnusuya SNo ratings yet
- ThalassaemiasDocument8 pagesThalassaemiasSalim UddinkhanNo ratings yet
- Thalasemia MedscapeDocument21 pagesThalasemia MedscapeFenty IswarNo ratings yet
- THALASSEMIADocument30 pagesTHALASSEMIAjismi vallachiraNo ratings yet
- Hematology Lectures 1 5 DR - TuyDocument10 pagesHematology Lectures 1 5 DR - TuyMiguel Cuevas DolotNo ratings yet
- Thalassaemias PDFDocument3 pagesThalassaemias PDFNidia100% (1)
- Hemoglobinopathies Occupy A Special Place in Human Genetics For Many ReasonsDocument17 pagesHemoglobinopathies Occupy A Special Place in Human Genetics For Many ReasonsjipmerbiochemNo ratings yet
- Pages From Download-2Document1 pagePages From Download-2Ra,i Al HaddadNo ratings yet
- Heme ChemistryDocument23 pagesHeme ChemistryShereen KaddafyNo ratings yet
- Hemoglobin Opa ThiesDocument34 pagesHemoglobin Opa ThiesFebri fitraNo ratings yet
- HemoglobinDocument7 pagesHemoglobinNinik Triayu S100% (1)
- A Guide To Life!: - and List To Live byDocument6 pagesA Guide To Life!: - and List To Live byBravan AliennNo ratings yet
- Computer Data Storage-1Document9 pagesComputer Data Storage-1Bravan AliennNo ratings yet
- 6 ThrombosisDocument27 pages6 ThrombosisBravan AliennNo ratings yet
- Vascularized Tissue To Injurious Agent A Protective ResponseDocument13 pagesVascularized Tissue To Injurious Agent A Protective ResponseBravan AliennNo ratings yet
- Cellular Events: A Critical Function of InflammationDocument21 pagesCellular Events: A Critical Function of InflammationBravan AliennNo ratings yet
- Bible Truth and The TruthDocument228 pagesBible Truth and The TruthJeff Stutz100% (1)
- Sickle Cell AnemiaDocument23 pagesSickle Cell AnemiaJesmin_36No ratings yet
- ThlasmiaDocument7 pagesThlasmiaSonia AhmedNo ratings yet
- ThalcemiaDocument33 pagesThalcemiaRahul Kumar DiwakarNo ratings yet
- Blood Typing ExperimentDocument6 pagesBlood Typing ExperimentAdib SadmanNo ratings yet
- Study On Hemoglobinopathies and G6PD Deficiency in Terai Districts of NepalDocument30 pagesStudy On Hemoglobinopathies and G6PD Deficiency in Terai Districts of NepalSaroj BhattaraiNo ratings yet
- Note That There Are Several Different "Basic Blast" Programs Available at Ncbi (Including Nucleotide Blast, Protein Blast, and Blastx)Document10 pagesNote That There Are Several Different "Basic Blast" Programs Available at Ncbi (Including Nucleotide Blast, Protein Blast, and Blastx)Raj Kumar SoniNo ratings yet
- Shivam Biology ProjectDocument18 pagesShivam Biology ProjectPankaj SolankiNo ratings yet
- Ann Druyan Talks About Carl Sagan and Science, Religion, Wonder, Awe - . .Document23 pagesAnn Druyan Talks About Carl Sagan and Science, Religion, Wonder, Awe - . .stephenking0078916No ratings yet
- AnemiaDocument33 pagesAnemiaJasmine Thabith100% (5)
- Case StudyDocument42 pagesCase StudyChristine Joy Ilao PasnoNo ratings yet
- Hereditary AnemiaDocument77 pagesHereditary AnemiaAravindh SivaNo ratings yet
- THALASSEMIADocument49 pagesTHALASSEMIAMunish DograNo ratings yet
- Army Public School Gopalpur: Chapter-5 Principles of Inheritance and Variation NotesDocument7 pagesArmy Public School Gopalpur: Chapter-5 Principles of Inheritance and Variation NotesAshok KumarNo ratings yet
- KVS Study Material Biology Class 12Document189 pagesKVS Study Material Biology Class 12Akshatha Nayak60% (5)
- Chapter 6 Hema 1Document7 pagesChapter 6 Hema 1EMAN SALEHNo ratings yet
- Sickle Cell AnemiaDocument37 pagesSickle Cell Anemiahazelposis75% (4)
- 3 ThalassemiaDocument20 pages3 ThalassemiaMARYLINENo ratings yet
- Dr. Silvanus Chakra PuspitaDocument74 pagesDr. Silvanus Chakra PuspitaDewi PratiwiNo ratings yet
- Structure, Types and Various Methods For Estimation of HaemoglobinDocument49 pagesStructure, Types and Various Methods For Estimation of HaemoglobinChandra ShekharNo ratings yet
- 12 Biology Revision Questions 2017 18 Chapter 5 PDFDocument15 pages12 Biology Revision Questions 2017 18 Chapter 5 PDFFaiz Alam100% (1)
- Bioinformatics-II (BIF501) : Assignment No.01Document15 pagesBioinformatics-II (BIF501) : Assignment No.01MehboobNo ratings yet
- DrepanocitozaDocument11 pagesDrepanocitozaElisa TelharajNo ratings yet
- AnemiaDocument145 pagesAnemiaTegegne WorkNo ratings yet
- Bioinformatic Analysis of HBBDocument1 pageBioinformatic Analysis of HBBfrank2689No ratings yet
- Background: Alpha ThalassemiaDocument24 pagesBackground: Alpha ThalassemiacristieristiieNo ratings yet
- 2 - Diagnosticul Molecular - Hype or HopeDocument57 pages2 - Diagnosticul Molecular - Hype or HopeSorin LazarNo ratings yet