Download as pdf or txt
You might also like
- Multiple Muscle Systems - Biomechanics and Movement OrganizationDocument817 pagesMultiple Muscle Systems - Biomechanics and Movement OrganizationChris SakeNo ratings yet
- Electric Welding Method Statement and Safe WorkDocument2 pagesElectric Welding Method Statement and Safe WorkBalumahendran Subramanian100% (1)
- Conditioning of AsilDocument8 pagesConditioning of AsilWorrenbuffet Rang60% (5)
- 4 - DS SATK Form - Transfer of Location 1.2Document3 pages4 - DS SATK Form - Transfer of Location 1.2Maybielyn DavidNo ratings yet
- Optimizing The Distribution of Leg Muscles For Vertical JumpingDocument15 pagesOptimizing The Distribution of Leg Muscles For Vertical JumpingWarren EnglishNo ratings yet
- Muscle Forces and The Demands of Human Walking: Adam D. Sylvester, Steven G. Lautzenheiser and Patricia Ann KramerDocument10 pagesMuscle Forces and The Demands of Human Walking: Adam D. Sylvester, Steven G. Lautzenheiser and Patricia Ann KramerCarmen Cuerdo del RíoNo ratings yet
- Warrener-2017 Hominin Hip Biomechanics - Changing PerspectivesDocument14 pagesWarrener-2017 Hominin Hip Biomechanics - Changing PerspectivesmaiNo ratings yet
- Crawford 2016Document25 pagesCrawford 2016walterNo ratings yet
- E128 Friday 17 October Papers / Journal of Science and Medicine in Sport 18S (2014) E108-E135Document1 pageE128 Friday 17 October Papers / Journal of Science and Medicine in Sport 18S (2014) E108-E135Jorge MenesesNo ratings yet
- OK 2015 McKeon - The Foot Core System - A New Paradigm For Understanding Intrinsic Foot Muscle FunctionDocument10 pagesOK 2015 McKeon - The Foot Core System - A New Paradigm For Understanding Intrinsic Foot Muscle FunctionMatias Santis OlivaresNo ratings yet
- Neuromechanical Interactions Between The Limbs DurDocument24 pagesNeuromechanical Interactions Between The Limbs DurWisnu Prasetyo AdhiNo ratings yet
- Predicting Gait Adaptations Due To Ankle Plantarflexor Muscle Weakness and Contracture Using Physics-Based Musculoskeletal SimulationsDocument27 pagesPredicting Gait Adaptations Due To Ankle Plantarflexor Muscle Weakness and Contracture Using Physics-Based Musculoskeletal Simulationskamran nazariNo ratings yet
- Ankle and Knee Extensor Muscle Effort During Locomotion in Young and Older Athletes: Implications For Understanding Age-Related Locomotor DeclineDocument7 pagesAnkle and Knee Extensor Muscle Effort During Locomotion in Young and Older Athletes: Implications For Understanding Age-Related Locomotor DeclineNguyễn HùngNo ratings yet
- Footcore SystemDocument10 pagesFootcore Systembarros6No ratings yet
- Damiano 2011 Comparisson Treadmill Walking CyclingDocument12 pagesDamiano 2011 Comparisson Treadmill Walking CyclingMauricio Alfredo Fuentes FloresNo ratings yet
- The Human Gluteus Maximus and Its Role in Running: CitationDocument14 pagesThe Human Gluteus Maximus and Its Role in Running: CitationIan ZhangNo ratings yet
- Tingle Et Al, 2023Document20 pagesTingle Et Al, 2023Livia PereiraNo ratings yet
- Journal of Biomechanics: Guillaume Christe, François Kade, Brigitte M. Jolles, Julien FavreDocument8 pagesJournal of Biomechanics: Guillaume Christe, François Kade, Brigitte M. Jolles, Julien FavreebrahimpanNo ratings yet
- 2015 - State of The Art in Prosthetic WristsDocument9 pages2015 - State of The Art in Prosthetic WristsMikiNo ratings yet
- Solution Manual For Anatomy Physiology The Unity of Form and Function 8th EditionDocument4 pagesSolution Manual For Anatomy Physiology The Unity of Form and Function 8th EditionJayStewartmsie100% (49)
- Dynamics of The Human Masticatory SystemDocument12 pagesDynamics of The Human Masticatory SystemLinda Garcia PNo ratings yet
- The Foot Core System. A New Paradigm For Understanding Intrinsic Foot Muscle FunctionDocument9 pagesThe Foot Core System. A New Paradigm For Understanding Intrinsic Foot Muscle FunctionVictor GonzálezNo ratings yet
- Powered Ankle Exoskeletons Existent Designs and Control SystemsDocument8 pagesPowered Ankle Exoskeletons Existent Designs and Control SystemsHeeralNo ratings yet
- The Foot Core System 2015Document10 pagesThe Foot Core System 2015ryankelsallNo ratings yet
- Current Trends in The Biokinetic Analysis of The Foot and AnkleDocument6 pagesCurrent Trends in The Biokinetic Analysis of The Foot and AnkleCarlos gutierrezNo ratings yet
- Applications of Upper Limb Biomechanical Models in Spinal Cord Injury PatientsDocument41 pagesApplications of Upper Limb Biomechanical Models in Spinal Cord Injury PatientsMarta Leal EscribanoNo ratings yet
- Biomechanics of Overground vs. Treadmill Walking in Healthy IndividualsDocument9 pagesBiomechanics of Overground vs. Treadmill Walking in Healthy IndividualsAlvaroNo ratings yet
- The Genetics of Human Performance: Daniel Seung Kim, Matthew T. Wheeler and Euan A. AshleyDocument15 pagesThe Genetics of Human Performance: Daniel Seung Kim, Matthew T. Wheeler and Euan A. AshleyThiago SartiNo ratings yet
- Hemiparetic Gait: Lynne R. Sheffler,, John ChaeDocument13 pagesHemiparetic Gait: Lynne R. Sheffler,, John ChaeassyifaNo ratings yet
- 5 Free Research ReviewsDocument20 pages5 Free Research ReviewsKelly AquinoNo ratings yet
- D H M S: Ynamics of The Uman Asticatory YstemDocument11 pagesD H M S: Ynamics of The Uman Asticatory YstemHugo A. Cárdenas IglesiasNo ratings yet
- Human KinesiologyDocument26 pagesHuman KinesiologyTrinidad Farías PiñeiroNo ratings yet
- Skeleton SystemDocument14 pagesSkeleton SystemDiscord YtNo ratings yet
- 2d70c0e7-a0d7-4d00-accb-a4697b7baaf1Document28 pages2d70c0e7-a0d7-4d00-accb-a4697b7baaf1ankit97bNo ratings yet
- Current Research in Physiology: Collin CarrollDocument8 pagesCurrent Research in Physiology: Collin CarrollAdrianSierradelReyNo ratings yet
- Human Movement Science: Brian C. Nairn, Janessa D.M. DrakeDocument11 pagesHuman Movement Science: Brian C. Nairn, Janessa D.M. DrakeQuiroprácticaParaTodosNo ratings yet
- A Control System For A Flexible Spine Belly-Dancing HumanoidDocument26 pagesA Control System For A Flexible Spine Belly-Dancing HumanoidAntonio SanNo ratings yet
- How Animals Move 3Document8 pagesHow Animals Move 3Juan Pablo Vargas GarciaNo ratings yet
- Effectiveness of Robotic Exoskeletons For Improving Gait in Children With Cerebral Palsy - A Systematic ReviewDocument12 pagesEffectiveness of Robotic Exoskeletons For Improving Gait in Children With Cerebral Palsy - A Systematic ReviewAkim KbNo ratings yet
- Dynamics of The Human Masticatory System. ReviewDocument12 pagesDynamics of The Human Masticatory System. Reviewjrr158No ratings yet
- Fisiologia Do Exercício Até Medalhas - 230828 - 113031Document96 pagesFisiologia Do Exercício Até Medalhas - 230828 - 113031peroNo ratings yet
- 4134 FullDocument6 pages4134 FullmaiNo ratings yet
- Anatomy and Physiology Research Paper TopicsDocument4 pagesAnatomy and Physiology Research Paper Topicsfzqs7g1d100% (1)
- GaitDocument8 pagesGaitPravin YadavNo ratings yet
- 2016 - Thinking, Walking, Talking - Integratory Motor and Cognitive Brain FunctionDocument19 pages2016 - Thinking, Walking, Talking - Integratory Motor and Cognitive Brain FunctionRaúl VerdugoNo ratings yet
- ARTIGO - Cazzola, Pavei, Preatoni - 2016 - Can Coordination Variability Identify Performance Factors and Skill Level in Competitive Sport The CasDocument9 pagesARTIGO - Cazzola, Pavei, Preatoni - 2016 - Can Coordination Variability Identify Performance Factors and Skill Level in Competitive Sport The CasKinetic LabNo ratings yet
- Variation of Rotation Moment Arms With Hip Flexion: Scott L. Delp, William E. Hess, David S. Hungerford, Lynne C. JonesDocument9 pagesVariation of Rotation Moment Arms With Hip Flexion: Scott L. Delp, William E. Hess, David S. Hungerford, Lynne C. JonesAryan SalihNo ratings yet
- Are Current Measurements of Lower Extremity MuscleDocument10 pagesAre Current Measurements of Lower Extremity MuscleLina M GarciaNo ratings yet
- Skeletal Muscle: A Brief Review of Structure and Function: Calcified Tissue International October 2014Document14 pagesSkeletal Muscle: A Brief Review of Structure and Function: Calcified Tissue International October 2014Anonymous K9tmd7T0PzNo ratings yet
- Journal of Neuroengineering and RehabilitationDocument16 pagesJournal of Neuroengineering and RehabilitationFikri DzakyNo ratings yet
- McKeon PO, Hertel J, Bramble D, & Davi I. The Foot Core System A New Paradigm For Understanding Intrinsic Foot Muscle Function PDFDocument9 pagesMcKeon PO, Hertel J, Bramble D, & Davi I. The Foot Core System A New Paradigm For Understanding Intrinsic Foot Muscle Function PDFLucas De MariaNo ratings yet
- 290 FullDocument9 pages290 Fulljovagu35No ratings yet
- PIIS2405844022003115Document10 pagesPIIS2405844022003115adr3an.tabaresNo ratings yet
- 9e10 PDFDocument19 pages9e10 PDFD'von AbiaNo ratings yet
- Bobbert Et Al., 2006 SJ - Explanation of The Bilateral Deficit in Human Vertical Squat JumpingDocument8 pagesBobbert Et Al., 2006 SJ - Explanation of The Bilateral Deficit in Human Vertical Squat JumpingIsmenia HelenaNo ratings yet
- Locomotion, Vertebrate: Auke Jan IjspeertDocument6 pagesLocomotion, Vertebrate: Auke Jan IjspeertAlissa QothrunnadaNo ratings yet
- Furrer Et Al 2023 the Molecular Athlete Exercise Physiology From Mechanisms to MedalsDocument96 pagesFurrer Et Al 2023 the Molecular Athlete Exercise Physiology From Mechanisms to MedalsMarcos CavalcanteNo ratings yet
- State-Of-The-Art Research in Robotic Hip Exoskeletons A General ReviewDocument10 pagesState-Of-The-Art Research in Robotic Hip Exoskeletons A General Reviewahmad joharaNo ratings yet
- Human Hip EvolutionDocument12 pagesHuman Hip EvolutionJeffersonNo ratings yet
- Maurel Et Al. - 2002 - A Biomechanical Musculoskeletal Model of Human Upper Limb For Dynamic SimulationDocument16 pagesMaurel Et Al. - 2002 - A Biomechanical Musculoskeletal Model of Human Upper Limb For Dynamic SimulationkoushikbakshiNo ratings yet
- The Effect of Stance On Muscle Activation in The Squat 1000 Word EssayDocument4 pagesThe Effect of Stance On Muscle Activation in The Squat 1000 Word Essayapi-254689954No ratings yet
- Advanced farriery knowledge: A study guide and AWCF theory course companionFrom EverandAdvanced farriery knowledge: A study guide and AWCF theory course companionNo ratings yet
- Machine Guarding StandardDocument68 pagesMachine Guarding StandardDave CNo ratings yet
- Shanghai Child and Adolescent Large-Scale Eye Study - High Myopia Registration Implementation PlanDocument27 pagesShanghai Child and Adolescent Large-Scale Eye Study - High Myopia Registration Implementation PlanPutra TridiyogaNo ratings yet
- II Epidemiology الكتابDocument76 pagesII Epidemiology الكتابAhmed ElhossanyNo ratings yet
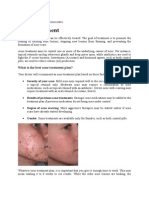
- Acne TreatmentDocument15 pagesAcne TreatmentSeptia Kurniaty100% (1)
- Formi NDF Chicken Broiler 4-Seiter Englisch Mit QR Code 08 2017 ScreenDocument4 pagesFormi NDF Chicken Broiler 4-Seiter Englisch Mit QR Code 08 2017 ScreenMichael MekhaNo ratings yet
- Jasmine Touch: Jasminum Grandiflorum 10 MLDocument1 pageJasmine Touch: Jasminum Grandiflorum 10 MLBeatrice LuanaNo ratings yet
- Developmental Dysplasia of The Hip (DDH)Document30 pagesDevelopmental Dysplasia of The Hip (DDH)whidiNo ratings yet
- Kickstart Your Time MNGMNT e 02Document5 pagesKickstart Your Time MNGMNT e 02Ngô TuấnNo ratings yet
- Waste ManagementDocument35 pagesWaste ManagementAnnabelle MañozoNo ratings yet
- Outcome of Bonded Vs All-Ceramic and PDFDocument21 pagesOutcome of Bonded Vs All-Ceramic and PDFsalshabilaNo ratings yet
- PathoConceptMap AIDSDocument3 pagesPathoConceptMap AIDSKristen Babauta50% (2)
- MEDI7112 GIT Lecture Checklist 2015Document2 pagesMEDI7112 GIT Lecture Checklist 2015zjaffri89No ratings yet
- 2003 Drug Induced Nightmares, An Etiology Based ReviewDocument9 pages2003 Drug Induced Nightmares, An Etiology Based ReviewDiego HormacheaNo ratings yet
- Vilas County News-Review, Nov. 16, 2011 - SECTION BDocument10 pagesVilas County News-Review, Nov. 16, 2011 - SECTION BNews-ReviewNo ratings yet
- Music Therapy and Eating DisordersDocument21 pagesMusic Therapy and Eating DisordersChristie McKernanNo ratings yet
- CE Courses Accredited: CE Department Celebrated Science.01Document8 pagesCE Courses Accredited: CE Department Celebrated Science.01kiwipangNo ratings yet
- Toddler DevelopmentDocument3 pagesToddler Developmentazooooz8m7376No ratings yet
- Shanda's Personal Strategic Plan 2018: Strategic Anchors Strategic Areas of FocusDocument1 pageShanda's Personal Strategic Plan 2018: Strategic Anchors Strategic Areas of Focusbirhanu Ayenew AnleyNo ratings yet
- Justificación PtarDocument3 pagesJustificación PtarAna Yulieth García GarcíaNo ratings yet
- JP-K107, 1107K: Print Date: 08.03.2018 Page 1 of 9Document9 pagesJP-K107, 1107K: Print Date: 08.03.2018 Page 1 of 9Junior BautistaNo ratings yet
- DiabetekDocument15 pagesDiabetekMaira GonzálezNo ratings yet
- MACD FormDocument44 pagesMACD FormNik ShNo ratings yet
- 4 - The Unsteady BookeeperDocument4 pages4 - The Unsteady Bookeeperkvguan7090No ratings yet
- Pediatric Otolaryngology For The Clinician 2009Document237 pagesPediatric Otolaryngology For The Clinician 2009Diaha100% (2)
- Physical Education and Health: Quarter I - Module 1Document25 pagesPhysical Education and Health: Quarter I - Module 1Gail Valdez100% (1)
- Kualitas Hidup Pasien Gagal Jantung Kongestif (GJK) Berdasarkan Karakteristik Demografi Arif Nur Akhmad, Yanuar Primanda, Yuni Permatasari IstantiDocument8 pagesKualitas Hidup Pasien Gagal Jantung Kongestif (GJK) Berdasarkan Karakteristik Demografi Arif Nur Akhmad, Yanuar Primanda, Yuni Permatasari IstantiHaji Mi'el Na Nur'ilmiNo ratings yet
- Placenta Previa (NCP)Document2 pagesPlacenta Previa (NCP)jonna casumpangNo ratings yet