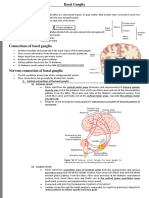
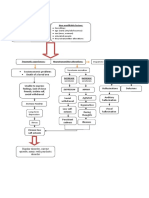
Download as pdf or txt
You might also like
- Babylon 5 RPG (1st Ed.) - The Minbari Federation Fact BookDocument202 pagesBabylon 5 RPG (1st Ed.) - The Minbari Federation Fact Booksunhawk88100% (6)
- Critical Thinking Development A Stage TheoryDocument12 pagesCritical Thinking Development A Stage Theoryconditioned_genesis1504No ratings yet
- ToeflDocument67 pagesToeflCseCseNo ratings yet
- Hand Functions in The Child 2006 PDFDocument476 pagesHand Functions in The Child 2006 PDFMaria Ramona100% (3)
- Space-Form and The Acousmatic Image - DENIS SMALLEYDocument25 pagesSpace-Form and The Acousmatic Image - DENIS SMALLEYLorena GaláxiaNo ratings yet
- Somatosensory PathwaysDocument36 pagesSomatosensory PathwaysAnonymous w3vYureNo ratings yet
- Symposium Neural PathwaysDocument27 pagesSymposium Neural PathwaysFasica MesfinNo ratings yet
- CerebellumDocument22 pagesCerebellumSalman XahirNo ratings yet
- CerebellumDocument51 pagesCerebellumLiya ThaaraNo ratings yet
- The Central Nervous System: Andi AnsharullahDocument60 pagesThe Central Nervous System: Andi AnsharullahAnonymous vUl83IptiSNo ratings yet
- Handouts MBBII ContinuedDocument18 pagesHandouts MBBII ContinuedLily KruegerNo ratings yet
- Special Senses Physiology of Hearing and Balance: Dr. CalaquianDocument6 pagesSpecial Senses Physiology of Hearing and Balance: Dr. CalaquianHanako Sasaki AranillaNo ratings yet
- The Neurological Exam Made SimpleDocument49 pagesThe Neurological Exam Made SimpleRizki AmaliahNo ratings yet
- Unit 3Document21 pagesUnit 3dekNo ratings yet
- Anatomy of Vestibular SystemDocument25 pagesAnatomy of Vestibular SystemFranciscus BuwanaNo ratings yet
- Late Onset Psychosis - A Case ReportDocument5 pagesLate Onset Psychosis - A Case ReportJAVED ATHER SIDDIQUINo ratings yet
- Sensory IntegrationDocument2 pagesSensory Integrationapi-165286810No ratings yet
- Cerebellar Disorders: Dr. Mohamed Nasreldin HamdoonDocument14 pagesCerebellar Disorders: Dr. Mohamed Nasreldin HamdoonMohamed Nasreldin HamdoonNo ratings yet
- Dopaminergic and Serotonergic Neurohumoural TransmissionDocument23 pagesDopaminergic and Serotonergic Neurohumoural TransmissionChaitanya Kiran PullelaNo ratings yet
- Convulsions (Seizures)Document26 pagesConvulsions (Seizures)Harmony OyikoNo ratings yet
- Gross Anatomy Learning Objectives - LimbsDocument8 pagesGross Anatomy Learning Objectives - Limbskep1313No ratings yet
- Is Depression With Increased Antithyroid Autoantibodies A Mere Chance Finding or Hashimoto's Encephalopathy PDFDocument2 pagesIs Depression With Increased Antithyroid Autoantibodies A Mere Chance Finding or Hashimoto's Encephalopathy PDFwilliams882No ratings yet
- Ascending Tracts Neuro AnatomyDocument2 pagesAscending Tracts Neuro AnatomyrachellesliedeleonNo ratings yet
- Motor TractDocument4 pagesMotor TractRaden Achmad Candra PutraNo ratings yet
- Jurnal AutismeDocument11 pagesJurnal AutismeKabhithra ThiayagarajanNo ratings yet
- Subject: Neurophysiology Topic: Hemispheric Organization Lecturer: Dr. Simbulan Date: March 2011Document12 pagesSubject: Neurophysiology Topic: Hemispheric Organization Lecturer: Dr. Simbulan Date: March 2011Std DlshsiNo ratings yet
- Anat 6.5 Basal Ganglia - QuijanoDocument6 pagesAnat 6.5 Basal Ganglia - Quijanolovelots1234No ratings yet
- Rtms Technique and Current Status: Dr. Shravan KumarDocument78 pagesRtms Technique and Current Status: Dr. Shravan KumarfitriaNo ratings yet
- Functional Neural CircuitsDocument1 pageFunctional Neural CircuitsUSCBISCNo ratings yet
- 9 PsychopathologyDocument2 pages9 PsychopathologyMonchee YusonNo ratings yet
- Toxic Metabolic Encephalopathy John W. Melton, M.DDocument17 pagesToxic Metabolic Encephalopathy John W. Melton, M.DabdalkhalidNo ratings yet
- Basal GangliaDocument6 pagesBasal Ganglia381a53c99bNo ratings yet
- Copy-Classification of Psychiatric DisordersDocument56 pagesCopy-Classification of Psychiatric Disordersshadrach bineyNo ratings yet
- Physiology FC PainDocument4 pagesPhysiology FC Painiqz saniomeNo ratings yet
- The Cerebelum 2Document53 pagesThe Cerebelum 2Reagan MwesigwaNo ratings yet
- Disorders of Thought and SpeechDocument50 pagesDisorders of Thought and SpeechTilarupa BhattaraiNo ratings yet
- The Descending Tracts of The Spinal CordDocument2 pagesThe Descending Tracts of The Spinal Cordalmastar officeNo ratings yet
- Chapter 5 Perception and ConsciousnessDocument5 pagesChapter 5 Perception and ConsciousnessAnn NguyenNo ratings yet
- 1028 - Temporal LobeDocument19 pages1028 - Temporal LobeEdward LinNo ratings yet
- Cerebellum and Basal Ganglia: Presented byDocument68 pagesCerebellum and Basal Ganglia: Presented byDivya AggarwalNo ratings yet
- Revised PathoDocument2 pagesRevised PathoFranz Earl Niño AlbesaNo ratings yet
- Neurohormonal Regulation of Body FunctionsDocument34 pagesNeurohormonal Regulation of Body FunctionsjimmyNo ratings yet
- Lecture 30 Histology of CnsDocument56 pagesLecture 30 Histology of CnsJustin DawsonNo ratings yet
- Moca Vs MMSEDocument2 pagesMoca Vs MMSEYw LiuNo ratings yet
- My Basal GangliaDocument18 pagesMy Basal GangliaPriya GKNo ratings yet
- Brain Anatomy, Anatomy of The Human BrainDocument12 pagesBrain Anatomy, Anatomy of The Human BrainElmir ƏzimovNo ratings yet
- Terms in The Field of PsychiatryDocument18 pagesTerms in The Field of PsychiatryOchie YecyecanNo ratings yet
- Fpga Implementation of Neural Networks: Main ContentsDocument21 pagesFpga Implementation of Neural Networks: Main Contentsatef BenhaouesNo ratings yet
- PRES (Posterior Reversible Encephalopathy Syndrome) and Eclampsia-ReviewDocument5 pagesPRES (Posterior Reversible Encephalopathy Syndrome) and Eclampsia-ReviewAvicena M IqbalNo ratings yet
- 18 - Thalamus and Limbic System (Edited)Document22 pages18 - Thalamus and Limbic System (Edited)Fotocopias LulisNo ratings yet
- Lect. 1 Organization of The Nervous System 54Document74 pagesLect. 1 Organization of The Nervous System 54nopporn2520No ratings yet
- Neurology Topics: Degenerative DiseasesDocument57 pagesNeurology Topics: Degenerative Diseasesलुकास विडालNo ratings yet
- Anat 6.6 Cerebellum - DeriadaDocument11 pagesAnat 6.6 Cerebellum - Deriadalovelots1234100% (1)
- CNS - Limbic SystemDocument46 pagesCNS - Limbic SystemsarvinaNo ratings yet
- Delirium Screening PRISMEDocument1 pageDelirium Screening PRISMEWardah Fauziah El SofwanNo ratings yet
- Hypothalamus and Pituitary GlandDocument39 pagesHypothalamus and Pituitary GlandGish KioiNo ratings yet
- Mapping Progressive Brain Structural Changes in Early Alzheimer's DiseaseDocument27 pagesMapping Progressive Brain Structural Changes in Early Alzheimer's Diseasetonylee24No ratings yet
- 2018 Bio Day 3Document21 pages2018 Bio Day 3api-391411195No ratings yet
- Meninges, Ventricles - CSF - Study GuideDocument3 pagesMeninges, Ventricles - CSF - Study Guideshivani patelNo ratings yet
- Cerebellar LesionsDocument3 pagesCerebellar LesionsMary100% (1)
- Psychiatry Kasr (2020-2021) @AUData - BotDocument119 pagesPsychiatry Kasr (2020-2021) @AUData - Botibrahim-t-abuawwadNo ratings yet
- Referred PainDocument3 pagesReferred PainaspalavistaNo ratings yet
- Anatomia Si Localizare in Boli SpinaleDocument8 pagesAnatomia Si Localizare in Boli SpinaleHalit DianaNo ratings yet
- Spinal Cord LesionsDocument110 pagesSpinal Cord Lesionsgtaha80100% (1)
- Environmental Biology Seminar PresentationDocument20 pagesEnvironmental Biology Seminar Presentationadedoyin opereNo ratings yet
- What Is A Physical Disability?: Spinal Cord Injury (SCI)Document14 pagesWhat Is A Physical Disability?: Spinal Cord Injury (SCI)XIARESSE DELA CRUZNo ratings yet
- Hemel Carmen - Using Haptic Imagery To Influence Online Product ExperiencesDocument34 pagesHemel Carmen - Using Haptic Imagery To Influence Online Product ExperiencesHappy NagashettiNo ratings yet
- Somatic Anatomy Body SystemsDocument7 pagesSomatic Anatomy Body Systemslomje l100% (1)
- Sensory ReeducationDocument52 pagesSensory Reeducationapi-373686592% (26)
- 1 s2.0 S096098221730708X MainDocument5 pages1 s2.0 S096098221730708X Mainrotinda bilekNo ratings yet
- Plan de Apoyo Inglés 1° Periodo 2021Document9 pagesPlan de Apoyo Inglés 1° Periodo 2021Mauricio RodriguezNo ratings yet
- HES 101 - Lesson 11-SensesDocument4 pagesHES 101 - Lesson 11-SensesLAGUERTA, JOHN MICHAEL A.No ratings yet
- Cerebrum Parietal Lobe Anatomy & Function AtfDocument2 pagesCerebrum Parietal Lobe Anatomy & Function AtfAdil Nawaz KhanNo ratings yet
- Benedict M Wand The Fit For Purpose ModelDocument13 pagesBenedict M Wand The Fit For Purpose ModelFirmansyah PurwantoNo ratings yet
- Concept Board Mood BoardDocument2 pagesConcept Board Mood BoardDiane MarananNo ratings yet
- Learning Styles InventoryDocument4 pagesLearning Styles InventoryQuanitum ExpressNo ratings yet
- ENGLISH 10 Lesson Plan A4 March 20, 2023 (Quarter 3)Document4 pagesENGLISH 10 Lesson Plan A4 March 20, 2023 (Quarter 3)Hannah ArguellesNo ratings yet
- Preterm Labor Case StudyDocument26 pagesPreterm Labor Case StudyKirk Espanol Bigstone82% (11)
- Food Choices of Tactile Defensive ChildrenDocument6 pagesFood Choices of Tactile Defensive ChildrenHilda RiveraNo ratings yet
- Pharmacy Catalog PDFDocument60 pagesPharmacy Catalog PDFDen IndiaNo ratings yet
- Physiology of Receptors Lec JPDocument55 pagesPhysiology of Receptors Lec JPabhishchheladiya0860No ratings yet
- CNS ExaminationDocument3 pagesCNS ExaminationsekharNo ratings yet
- B. Caseau, Tactile Piety in Knowing Bodies 2017Document32 pagesB. Caseau, Tactile Piety in Knowing Bodies 2017beatrice chevallier caseauNo ratings yet
- An Algorithm For The Evaluation of Peripheral Neuropathy - American Family PhysicianDocument9 pagesAn Algorithm For The Evaluation of Peripheral Neuropathy - American Family PhysicianAnonymous vnv6QFNo ratings yet
- Restoring The Human TouchDocument6 pagesRestoring The Human Touchlimber dameciousNo ratings yet
- Male Seadxual Disfunction PDFDocument1,013 pagesMale Seadxual Disfunction PDFAlexandr TrotskyNo ratings yet
- Queer Phenomenology Sara AhmedDocument33 pagesQueer Phenomenology Sara AhmedGregory Gibson50% (2)
- Reading IeltsDocument5 pagesReading IeltsjohnlycasNo ratings yet
- Print Chapter 13 Flashcards - Easy NotecardsDocument18 pagesPrint Chapter 13 Flashcards - Easy NotecardsoyamaNo ratings yet
- SEQs Key Send-Up 2nd Year Mbbs Amna Inayat Medical CollegeDocument16 pagesSEQs Key Send-Up 2nd Year Mbbs Amna Inayat Medical CollegeMudassar Roomi100% (1)