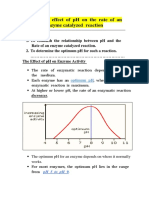
Download as pdf or txt
You might also like
- ENK Enzyme Kinetics Lab ManualDocument11 pagesENK Enzyme Kinetics Lab ManualVanesha AnesNo ratings yet
- Enzyme AP LabDocument8 pagesEnzyme AP LabMimiNo ratings yet
- Advanced Pharmaceutical analysisFrom EverandAdvanced Pharmaceutical analysisRating: 4.5 out of 5 stars4.5/5 (2)
- ENZYMES COMPLETE NOTES (UNIT 5 - B.Pharm 2nd Sem) PDFDocument15 pagesENZYMES COMPLETE NOTES (UNIT 5 - B.Pharm 2nd Sem) PDFBhavana Gangurde92% (36)
- Catalase AEBIDocument6 pagesCatalase AEBIGabriel CamarenaNo ratings yet
- A Simplification of The Protein Assay Method of Lowry Et Al. Which Is More Generally Applicable PDFDocument11 pagesA Simplification of The Protein Assay Method of Lowry Et Al. Which Is More Generally Applicable PDFTania SandovalNo ratings yet
- Proceedings The National: of Academy of SciencesDocument6 pagesProceedings The National: of Academy of SciencesSochenda PenNo ratings yet
- Catalase: Hans LuckDocument10 pagesCatalase: Hans LuckNguyễn QuangNo ratings yet
- 1971 R K Kulkarni E G Moore A F Hegyeli - Fred Leonard (1971) - Biodegradable Poly (Lactic Acid) Polymers 5 (3) 169-181 1971Document13 pages1971 R K Kulkarni E G Moore A F Hegyeli - Fred Leonard (1971) - Biodegradable Poly (Lactic Acid) Polymers 5 (3) 169-181 1971Julio ArruaNo ratings yet
- Efectos de Las Concentraciones de Oxígeno Disuelto y Ácido Acético en La Producción de Ácido Acético en El Cultivo Continuo de Acetobacter AcetiDocument6 pagesEfectos de Las Concentraciones de Oxígeno Disuelto y Ácido Acético en La Producción de Ácido Acético en El Cultivo Continuo de Acetobacter AcetiWendy Rivera100% (1)
- IsotachophoresisDocument7 pagesIsotachophoresisEka HerlinaNo ratings yet
- Dendrimetos de PTDocument16 pagesDendrimetos de PTJose Miguel PalomoNo ratings yet
- A Spectrophotometric Method For Determination of Catalase Activity in Small Tissue SamplesDocument6 pagesA Spectrophotometric Method For Determination of Catalase Activity in Small Tissue Samplesreza rezaieNo ratings yet
- Exp 2-Starch Hydrolysis by AmylaseDocument6 pagesExp 2-Starch Hydrolysis by AmylaseDodi Ihsan TaufiqNo ratings yet
- Aldol Puzzle Lab ReportDocument11 pagesAldol Puzzle Lab Reportlaurabruce27100% (1)
- Biological Activity and Variation of Toxicity of Probenecid-IJACS-2018Document7 pagesBiological Activity and Variation of Toxicity of Probenecid-IJACS-2018Soumen BhattacharjeeNo ratings yet
- Effect of PH On Enzyme ActivityDocument12 pagesEffect of PH On Enzyme ActivityAb AbNo ratings yet
- Exp 2-Starch Hydrolysis by AmylaseDocument6 pagesExp 2-Starch Hydrolysis by AmylaseGintoki SakataNo ratings yet
- 3 - Manole 8pagDocument8 pages3 - Manole 8pagVincent MalayaoNo ratings yet
- H NMR Study of The Effectiveness of Various Thiols For Removal of Methylmercury From Hemolyzed ErythrocytesDocument11 pagesH NMR Study of The Effectiveness of Various Thiols For Removal of Methylmercury From Hemolyzed ErythrocytesWalid EbaiedNo ratings yet
- Selulosa 4Document12 pagesSelulosa 4Luna Purnama Apriani SilaenNo ratings yet
- To Whom Reprint Requests Should Be AddressedDocument11 pagesTo Whom Reprint Requests Should Be AddressedAracely FloresNo ratings yet
- Inhibition of Tetrahydrolipstatin: Pancreatic inDocument5 pagesInhibition of Tetrahydrolipstatin: Pancreatic inPatrick Del PradoNo ratings yet
- Organic Chemistry III Laboratory: NMR Verification of Diastereoselective Reduction of Substituted CyclohexanonesDocument4 pagesOrganic Chemistry III Laboratory: NMR Verification of Diastereoselective Reduction of Substituted Cyclohexanonesungu_sakuraNo ratings yet
- Research Article: Rapid Elimination of Blood Alcohol Using Erythrocytes: Mathematical Modeling and in Vitro StudyDocument19 pagesResearch Article: Rapid Elimination of Blood Alcohol Using Erythrocytes: Mathematical Modeling and in Vitro StudyCristian BenalcázarNo ratings yet
- Hydrogen Peroxide DecompositionDocument17 pagesHydrogen Peroxide DecompositionChristian Eduardo Fabian50% (2)
- Mladen Oska 2007Document5 pagesMladen Oska 2007Shikhin GargNo ratings yet
- Reaction Rate and Activation Energy of The Acidolysis of Ethyl AcetateDocument6 pagesReaction Rate and Activation Energy of The Acidolysis of Ethyl AcetateGlënn Märk PrësörësNo ratings yet
- Esterification Process To Synthesize Isopropyl Chloroacetate Catalyzed by Lanthanum Dodecyl SulfateDocument6 pagesEsterification Process To Synthesize Isopropyl Chloroacetate Catalyzed by Lanthanum Dodecyl SulfateVinay JainNo ratings yet
- Rice Hull To Kojic AcidDocument17 pagesRice Hull To Kojic AcidArrianne Jaye MataNo ratings yet
- Lott 1975 Evaluation of Trinder's Glucose Oxidase Method For Measuring Glucose in Serunm An D UrineDocument7 pagesLott 1975 Evaluation of Trinder's Glucose Oxidase Method For Measuring Glucose in Serunm An D UrineАксель ЛюдовикичNo ratings yet
- Transactions of Tianjin University Volume 14 Issue 1 2008Document6 pagesTransactions of Tianjin University Volume 14 Issue 1 2008Sagar DhuriNo ratings yet
- Iodometric Determination of GlucoseDocument2 pagesIodometric Determination of GlucoseAnuj jainNo ratings yet
- Storer RJ Ferrante A 1998 Meth Mol Biol 79 81-90 chp:10.1385/0-89603-448-8:81Document10 pagesStorer RJ Ferrante A 1998 Meth Mol Biol 79 81-90 chp:10.1385/0-89603-448-8:81rjstorer9No ratings yet
- Laboratory Analytical Procedure: Chemical Analysis and Testing TaskDocument8 pagesLaboratory Analytical Procedure: Chemical Analysis and Testing TaskJuan Alzate JaramilloNo ratings yet
- Modelling Ethanol Production From Cellulose: Separate Hydrolysis and Fermentation Versus Simultaneous Saccharification and FermentationDocument10 pagesModelling Ethanol Production From Cellulose: Separate Hydrolysis and Fermentation Versus Simultaneous Saccharification and FermentationlandolandazNo ratings yet
- Biochemical Properties of A B-Xylosidase From: Clostridium CellulolyticumDocument4 pagesBiochemical Properties of A B-Xylosidase From: Clostridium CellulolyticumPedro HamannNo ratings yet
- PFR Lab ReportDocument16 pagesPFR Lab Reportcog0812No ratings yet
- Electrochemical Bienzyme Membrane Sensor For Free CholesterolDocument11 pagesElectrochemical Bienzyme Membrane Sensor For Free CholesterolAnand ChelliahNo ratings yet
- Mechanism of Cytoplasmatic PH Regulation in Hypoxic Maize Root Tips and Its Role in Survival Under HypoxiaDocument5 pagesMechanism of Cytoplasmatic PH Regulation in Hypoxic Maize Root Tips and Its Role in Survival Under HypoxiaGabriel ZahariaNo ratings yet
- Experiment 1 Ester SapnificationDocument14 pagesExperiment 1 Ester SapnificationTajTaj100% (1)
- Antioxidant EnzymesDocument20 pagesAntioxidant EnzymeselhadyegNo ratings yet
- 1969 - The Redox State of Free Nicotinamide-Adenine Dinucleotide Phosphate in The Cytoplasm of Rat LiverDocument11 pages1969 - The Redox State of Free Nicotinamide-Adenine Dinucleotide Phosphate in The Cytoplasm of Rat LiverRaymond LaBoyNo ratings yet
- Hydrogen Peroxide DecompositionDocument17 pagesHydrogen Peroxide DecompositionTan Xin YuNo ratings yet
- Simulation of Enzymatic Hydrolysis of Wheat Straw Using Autohydrolysis and OrganosolvDocument1 pageSimulation of Enzymatic Hydrolysis of Wheat Straw Using Autohydrolysis and OrganosolvSemana da Escola de Engenharia da Universidade do MinhoNo ratings yet
- Liquid-Liquid Separation in Solutions of Normal and Sickle Cell HemoglobinDocument5 pagesLiquid-Liquid Separation in Solutions of Normal and Sickle Cell HemoglobinFadi Simon de Souza MagalhãesNo ratings yet
- tmpF0F1 TMPDocument11 pagestmpF0F1 TMPFrontiersNo ratings yet
- Kinetics and Mechanisms of Antioxidant Activity Using The DPPH Free Radical MethodDocument7 pagesKinetics and Mechanisms of Antioxidant Activity Using The DPPH Free Radical Methodseema yadavNo ratings yet
- 1 s2.0 S0021925818559421 MainDocument15 pages1 s2.0 S0021925818559421 MainDat VuNo ratings yet
- Comparison of Acetylcholinesterase by Michel and Ellman MethodsDocument2 pagesComparison of Acetylcholinesterase by Michel and Ellman MethodschanNo ratings yet
- 22Document5 pages22AndresCuevasDiazNo ratings yet
- Ref (Ellman - 1961)Document9 pagesRef (Ellman - 1961)Tasso SalesNo ratings yet
- Question Bank: Metabolite Concentration ( - )Document2 pagesQuestion Bank: Metabolite Concentration ( - )Kanupriya TiwariNo ratings yet
- Enzyme Kinetics - Lab Manual 02-16-09 HMDocument10 pagesEnzyme Kinetics - Lab Manual 02-16-09 HMFoudiKarimNo ratings yet
- Free Radical and Antioxidant Protocols - Chapter 5Document6 pagesFree Radical and Antioxidant Protocols - Chapter 5Newocean NguyenNo ratings yet
- LT2 - Modelo Cinetico Pseudo-Intrinseco de La Bioconversion de CetonaDocument9 pagesLT2 - Modelo Cinetico Pseudo-Intrinseco de La Bioconversion de CetonaMafer CVNo ratings yet
- Liquid ALT (SGPT) Reagent SetDocument2 pagesLiquid ALT (SGPT) Reagent SetYahya RizkiNo ratings yet
- MMP-1 Practical Handout v1.0Document5 pagesMMP-1 Practical Handout v1.0Maisha JashimNo ratings yet
- White 1976Document10 pagesWhite 1976poojajain86No ratings yet
- Communication: Cuo/Sio: A Simple and Efficient Solid Acid Catalyst For Epoxide Ring OpeningDocument4 pagesCommunication: Cuo/Sio: A Simple and Efficient Solid Acid Catalyst For Epoxide Ring Openingfedericasantoro81No ratings yet
- N-Acyl Homoserine Lactone Analog Modulators of The Pseudomonas Aeruginosa RhII Quorum Signal Synthase, Shin Et Al.Document77 pagesN-Acyl Homoserine Lactone Analog Modulators of The Pseudomonas Aeruginosa RhII Quorum Signal Synthase, Shin Et Al.deryhermawanNo ratings yet
- Transition Metal Catalyzed Furans Synthesis: Transition Metal Catalyzed Heterocycle Synthesis SeriesFrom EverandTransition Metal Catalyzed Furans Synthesis: Transition Metal Catalyzed Heterocycle Synthesis SeriesNo ratings yet
- BBPK MCQ PciDocument3 pagesBBPK MCQ PcirajeshNo ratings yet
- WJPR Shivaleela Rev ArticleDocument26 pagesWJPR Shivaleela Rev ArticlerajeshNo ratings yet
- 3.6 Pharmaceutical Formulations (Theory)Document2 pages3.6 Pharmaceutical Formulations (Theory)rajeshNo ratings yet
- Formulation of Emulsions: Marie Wahlgren, Björn Bergenståhl, Lars Nilsson, and Marilyn RaynerDocument55 pagesFormulation of Emulsions: Marie Wahlgren, Björn Bergenståhl, Lars Nilsson, and Marilyn RaynerrajeshNo ratings yet
- JNTUH Revised Academic Calendar 2020-21 For B.Tech. and B.Pharm. II, III & IV Years I & II SemestersDocument1 pageJNTUH Revised Academic Calendar 2020-21 For B.Tech. and B.Pharm. II, III & IV Years I & II SemestersrajeshNo ratings yet
- I ST Pharm.D Remedial MathsDocument1 pageI ST Pharm.D Remedial MathsrajeshNo ratings yet
- 5.3 Clinical Pharmacokinetics and Pharmacotherapeutic Drug Monitoring (Theory)Document1 page5.3 Clinical Pharmacokinetics and Pharmacotherapeutic Drug Monitoring (Theory)rajesh100% (1)
- B.Pharmacy IY - R17 - Remedial Maths PDFDocument2 pagesB.Pharmacy IY - R17 - Remedial Maths PDFrajeshNo ratings yet
- Circular To Dental CollegesDocument3 pagesCircular To Dental CollegesrajeshNo ratings yet
- 4pain ManagementDocument1 page4pain ManagementrajeshNo ratings yet
- Functional Materials in Food NanotechnologyDocument10 pagesFunctional Materials in Food NanotechnologyrajeshNo ratings yet
- B.Pharmacy IY - R17 - Remedial MathsDocument2 pagesB.Pharmacy IY - R17 - Remedial MathsrajeshNo ratings yet
- Gaumet2008 PDFDocument9 pagesGaumet2008 PDFrajeshNo ratings yet
- SyllabusDocument107 pagesSyllabusrajeshNo ratings yet
- Population Pharmacokinetics II: Estimation MethodsDocument9 pagesPopulation Pharmacokinetics II: Estimation MethodsrajeshNo ratings yet
- Regulatory Issues & Addiction: Case SnippetDocument1 pageRegulatory Issues & Addiction: Case SnippetrajeshNo ratings yet
- Statistical Graphics in Pharmacokinetics and Pharmacodynamics: A TutorialDocument11 pagesStatistical Graphics in Pharmacokinetics and Pharmacodynamics: A TutorialrajeshNo ratings yet
- European Journal of Pharmaceutics and Biopharmaceutics: Yanzhuo Zhang, Tongying Jiang, Qiang Zhang, Siling WangDocument7 pagesEuropean Journal of Pharmaceutics and Biopharmaceutics: Yanzhuo Zhang, Tongying Jiang, Qiang Zhang, Siling WangrajeshNo ratings yet
- SUYAO WS1 Enzyme KineticsDocument8 pagesSUYAO WS1 Enzyme KineticsR SuyaoNo ratings yet
- Brahmadeo Dewprashad - The Chemistry of CocaineDocument8 pagesBrahmadeo Dewprashad - The Chemistry of CocainePoloGreenNo ratings yet
- Enzymes in Drug DiscoveryDocument8 pagesEnzymes in Drug Discoverypmpbzcbr8rNo ratings yet
- 7.016 Recitation 3 - Fall 2018: (Note: The Recitation Summary Should NOT Be Regarded As The Substitute For Lectures)Document6 pages7.016 Recitation 3 - Fall 2018: (Note: The Recitation Summary Should NOT Be Regarded As The Substitute For Lectures)Manish SarkarNo ratings yet
- Effect of Temperature On Enzyme Kinetics StudyDocument7 pagesEffect of Temperature On Enzyme Kinetics StudyYvonne MunNo ratings yet
- PSSHDocument7 pagesPSSHSaki ShaikhNo ratings yet
- Fermentation Kinetics For The Production of Ethanol by ImmobilizeDocument278 pagesFermentation Kinetics For The Production of Ethanol by ImmobilizeFitria Nur LailyNo ratings yet
- Enzyme Inhibition/Enzyme InhibitorsDocument5 pagesEnzyme Inhibition/Enzyme InhibitorsFaria bukhariNo ratings yet
- Purich 2010Document1 pagePurich 2010Stjepan BičanićNo ratings yet
- Enzyme Kinetics ExptsDocument13 pagesEnzyme Kinetics ExptsChemistryIndianguyNo ratings yet
- Toaz - Info Vet Mcqs PRDocument133 pagesToaz - Info Vet Mcqs PRYusra KhalidNo ratings yet
- Full Ebook of Novel Food Grade Enzymes Applications in Food Processing and Preservation Industries Abhishek Dutt Tripathi Kianoush Khosravi Darani Suresh Kumar Srivastava Eds Online PDF All ChapterDocument70 pagesFull Ebook of Novel Food Grade Enzymes Applications in Food Processing and Preservation Industries Abhishek Dutt Tripathi Kianoush Khosravi Darani Suresh Kumar Srivastava Eds Online PDF All Chapterlorenzobarta66778100% (7)
- Tyrosinase Enzyme InhibitorDocument8 pagesTyrosinase Enzyme Inhibitordamian_marie7769No ratings yet
- Comms 10008Document12 pagesComms 10008Hina MaqsoodNo ratings yet
- "Blue" LaccasesDocument15 pages"Blue" LaccasesVincent MalayaoNo ratings yet
- LineweaverBurk PlotDocument17 pagesLineweaverBurk PlotJawadNo ratings yet
- 5 FermentationDocument125 pages5 FermentationAkshitaNo ratings yet
- Kinetic Analysis of Tyrosinase Enzyme: Experiment #5Document39 pagesKinetic Analysis of Tyrosinase Enzyme: Experiment #5CareyTranNo ratings yet
- A Bacterium That Degrades and Assimilates Poly (Ethylene Terephthalate)Document5 pagesA Bacterium That Degrades and Assimilates Poly (Ethylene Terephthalate)Devita AmeliaNo ratings yet
- Section 8.5 Enzymes Can Be Inhibited by Specific MoleculesDocument8 pagesSection 8.5 Enzymes Can Be Inhibited by Specific MoleculesMohamed FaizalNo ratings yet
- Enzymes: Jason Ryan, MD, MPHDocument102 pagesEnzymes: Jason Ryan, MD, MPHMohamed AdelNo ratings yet
- Chemical+ +syllabus+7 8+SEM+175+Credit 2022 23Document50 pagesChemical+ +syllabus+7 8+SEM+175+Credit 2022 23d mNo ratings yet
- Palaganas Hw2 Che 197Document5 pagesPalaganas Hw2 Che 197Elah Palaganas100% (1)
- Enzyme: Oleh Yana Cahyana Stp.,Dea.,Ph.DDocument21 pagesEnzyme: Oleh Yana Cahyana Stp.,Dea.,Ph.DRina AryantiNo ratings yet
- A Generalised Enzyme Kinetic Model For Predicting The Behaviour of Complex Biochemical Systems PDFDocument14 pagesA Generalised Enzyme Kinetic Model For Predicting The Behaviour of Complex Biochemical Systems PDFEmanuel PataquivaNo ratings yet
- Enzyme KineticsDocument13 pagesEnzyme KineticsMohib100% (1)
- SM ch3 Bio Shuler 3Document19 pagesSM ch3 Bio Shuler 3Oluwaseun OyekolaNo ratings yet
- Non-Linear Pharmacokinetics: Biopharmaceutics (Ashik Sir)Document9 pagesNon-Linear Pharmacokinetics: Biopharmaceutics (Ashik Sir)Hasan Al MasudNo ratings yet