Download as pdf or txt
You might also like
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5822)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1093)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (852)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (590)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (898)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (540)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (349)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (823)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (122)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (403)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- Immunology MCQDocument50 pagesImmunology MCQAbdallah Essam Al-Zireeni96% (139)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (74)
- Quiz ImmunologyDocument50 pagesQuiz ImmunologyMedShare97% (39)
- Odors of Santity Distinctions of The Holy in Early Christianity and IslamDocument13 pagesOdors of Santity Distinctions of The Holy in Early Christianity and Islamkiedd_04100% (1)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- Cell PhysiologyDocument61 pagesCell Physiologykiedd_04100% (4)
- Microbiology Midterm ExamDocument8 pagesMicrobiology Midterm Examslg23350% (2)
- From The Bodies of Bees Classical and Christian Echoes in Surah Al-NahlDocument25 pagesFrom The Bodies of Bees Classical and Christian Echoes in Surah Al-Nahlkiedd_04No ratings yet
- Iklan Jabatan Pengajian Tinggi (Permohonan Kemasukan Tevt & Ilka) Sesi 2011/2012Document2 pagesIklan Jabatan Pengajian Tinggi (Permohonan Kemasukan Tevt & Ilka) Sesi 2011/2012kiedd_04No ratings yet
- "Traditional" Exegeses of Q 4:34Document15 pages"Traditional" Exegeses of Q 4:34kiedd_04No ratings yet
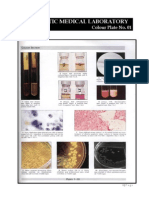
- Microbiology Colour Plate No.8Document1 pageMicrobiology Colour Plate No.8kiedd_04No ratings yet
- The Place For Others in IslamDocument27 pagesThe Place For Others in Islamkiedd_04No ratings yet
- Fadhilat Surah at TakwirDocument1 pageFadhilat Surah at Takwirkiedd_04No ratings yet
- Refleks ArcsDocument34 pagesRefleks Arcskiedd_04100% (1)
- Microbiology Colour Plate No.9Document1 pageMicrobiology Colour Plate No.9kiedd_04No ratings yet
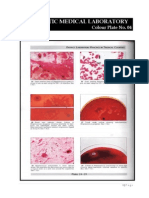
- Microbiology Colour Plate No.5Document1 pageMicrobiology Colour Plate No.5kiedd_04No ratings yet
- SYNAPSEDocument35 pagesSYNAPSEkiedd_04100% (3)
- Microbiology Colour Plate No.7Document1 pageMicrobiology Colour Plate No.7kiedd_04No ratings yet
- Microbiology Colour Plate No.6Document1 pageMicrobiology Colour Plate No.6kiedd_04No ratings yet
- Microbiology Colour Plate No.1Document1 pageMicrobiology Colour Plate No.1kiedd_04No ratings yet
- Microbiology Colour Plate No.3Document1 pageMicrobiology Colour Plate No.3kiedd_04No ratings yet
- Microbiology Colour Plate No.2Document1 pageMicrobiology Colour Plate No.2kiedd_04No ratings yet
- Microbiology Colour Plate No.4Document1 pageMicrobiology Colour Plate No.4kiedd_04No ratings yet
- Mitigation Potential and Costs Land-Use OptionsDocument9 pagesMitigation Potential and Costs Land-Use Optionskiedd_04No ratings yet
- Philosophical Consolation in Christianity and Islam Boethious and Al-KindiDocument10 pagesPhilosophical Consolation in Christianity and Islam Boethious and Al-Kindikiedd_04No ratings yet
- Stool Sample ProcessingDocument1 pageStool Sample Processingkiedd_04No ratings yet
- API® CoryneDocument4 pagesAPI® Corynekiedd_04No ratings yet
- Mitigation of Climate ChangeDocument41 pagesMitigation of Climate Changekiedd_04No ratings yet
- Oxidase TestDocument1 pageOxidase Testkiedd_04100% (1)
- Advance Diagnostic Medical Laboratory: The AP 20E® For Identification of BacteriaDocument1 pageAdvance Diagnostic Medical Laboratory: The AP 20E® For Identification of Bacteriakiedd_04100% (1)
- Mitigation of Climate ChangeDocument25 pagesMitigation of Climate Changekiedd_04No ratings yet
- Diagram of Classification of EnterobacteriaDocument1 pageDiagram of Classification of Enterobacteriakiedd_04100% (1)
- Carnitine DeficiencyDocument21 pagesCarnitine Deficiencykiedd_04100% (1)
- VMC Unit IV Notes 2017 BVSc&AH March 20 2019 PDFDocument99 pagesVMC Unit IV Notes 2017 BVSc&AH March 20 2019 PDFAbhijith S. PNo ratings yet
- Fimmu 10 01787Document13 pagesFimmu 10 01787Faiz SheeNo ratings yet
- PDF Middletons Allergy Essentials 1E 1 Har PSC Edition Robyn E Ohehir Fracp PHD Frcpath Ebook Full ChapterDocument63 pagesPDF Middletons Allergy Essentials 1E 1 Har PSC Edition Robyn E Ohehir Fracp PHD Frcpath Ebook Full Chapterkathy.cargle605100% (4)
- The NF-KB PathwayDocument4 pagesThe NF-KB PathwayAndri Praja Satria100% (1)
- B Cell Development 5th July 2013Document30 pagesB Cell Development 5th July 2013Rajkishor YadavNo ratings yet
- WORKSHEET 3 Lymphocyte ActivationDocument5 pagesWORKSHEET 3 Lymphocyte ActivationNeha ChoudharyNo ratings yet
- Introductory Immunology - Basic Concepts For Interdisciplinary Applications, 3e (Feb 6, 2023) - (0443153086) - (Academic Press) 3rd Edition ActorDocument43 pagesIntroductory Immunology - Basic Concepts For Interdisciplinary Applications, 3e (Feb 6, 2023) - (0443153086) - (Academic Press) 3rd Edition Actorwilliam.vickers704100% (9)
- 43 Lecture PresentationDocument113 pages43 Lecture Presentationashleymatrin123No ratings yet
- Autoimmune Bullous DiseasesDocument215 pagesAutoimmune Bullous Diseasessri karuniaNo ratings yet
- ImmunologyDocument55 pagesImmunologyW.F KareemNo ratings yet
- Advances in Immunology Vol. 111. Frederick W. Alt (Eds.)Document213 pagesAdvances in Immunology Vol. 111. Frederick W. Alt (Eds.)myjesa07No ratings yet
- Immune Response Regulation by Antigen Receptors' Clone-Specific Nonself PartsDocument14 pagesImmune Response Regulation by Antigen Receptors' Clone-Specific Nonself PartsMohammed R.HusseinNo ratings yet
- B CellDocument13 pagesB CellSrini VasanNo ratings yet
- Role of Rac1-Pak Pathway in Aggressive B-Cell LymphomaDocument169 pagesRole of Rac1-Pak Pathway in Aggressive B-Cell LymphomaShahid ShaikhNo ratings yet
- ImunologyDocument118 pagesImunologydzedalNo ratings yet
- Carlberg C. Molecular Immunology. How Science Works 2022Document228 pagesCarlberg C. Molecular Immunology. How Science Works 2022Philip Rodríguez GarcíaNo ratings yet
- Microbiology 1.1 Immunology - Dr. FontanillaDocument115 pagesMicrobiology 1.1 Immunology - Dr. FontanillaRyn ShadowNo ratings yet
- Ebook Autoimmune Diseases in Domestic Animals PDF Full Chapter PDFDocument67 pagesEbook Autoimmune Diseases in Domestic Animals PDF Full Chapter PDFsean.newport169100% (33)
- Immunology Last Year Final Exam 2011Document12 pagesImmunology Last Year Final Exam 2011Abdallah Essam Al-Zireeni80% (5)
- Cells of Immune System Notes 2Document70 pagesCells of Immune System Notes 2Sudeeksha Ravikoti100% (1)
- Systems Immunology An Introduction To Modeling MethodsDocument355 pagesSystems Immunology An Introduction To Modeling MethodsABDALSLAM ALNOURNo ratings yet
- Antigen and Antibody InteractionDocument17 pagesAntigen and Antibody InteractionSaumya PothukuchiNo ratings yet
- Multiple Myeloma - A Quick Reflection On The Fast Progress 2013Document326 pagesMultiple Myeloma - A Quick Reflection On The Fast Progress 2013Daoud IssaNo ratings yet
- Full Download Book Middletons Allergy Essentials 1E PDFDocument41 pagesFull Download Book Middletons Allergy Essentials 1E PDFbarry.schwend504100% (22)
- Immunogenetics - B Cell DevelopmentDocument38 pagesImmunogenetics - B Cell DevelopmentMadhu Mitha100% (1)
- Paul-Peter Tak-New Therapeutic Targets in Rheumatoid Arthritis (Progress in Inflammation Research) - Birkhäuser Basel (2009)Document233 pagesPaul-Peter Tak-New Therapeutic Targets in Rheumatoid Arthritis (Progress in Inflammation Research) - Birkhäuser Basel (2009)Subhadip BanerjeeNo ratings yet
- B Cells in Immunity and Tolerance - BookDocument190 pagesB Cells in Immunity and Tolerance - BookJuan FranciscoNo ratings yet