Download as pdf or txt
You might also like
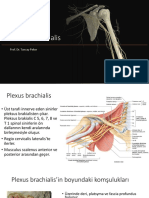
- Plexus BrachialisDocument32 pagesPlexus BrachialisunejsaNo ratings yet
- Tıbbi Biyolojiye Giriş Ve Hücre Biyolojisinin Kilometre TaşlarıDocument25 pagesTıbbi Biyolojiye Giriş Ve Hücre Biyolojisinin Kilometre Taşlarıjg7yr2978yNo ratings yet
- MitokondrionDocument31 pagesMitokondrionozkucuk10641No ratings yet
- HAFTA-Hücre KimyasìDocument32 pagesHAFTA-Hücre Kimyasìzy.shitsiteNo ratings yet
- A.YerlikayaMolekülerBiyoloji2011 (Sy 68 Den Başla) PDFDocument374 pagesA.YerlikayaMolekülerBiyoloji2011 (Sy 68 Den Başla) PDFÖzlem ErdemNo ratings yet
- Genetikte Temel KavramlarDocument187 pagesGenetikte Temel KavramlarkonuanlatimiNo ratings yet
- 9.sinif Biyoloji Hucre Konu AnlatimiDocument37 pages9.sinif Biyoloji Hucre Konu Anlatimimerve tanNo ratings yet
- Tıbbi Biyolojinin Tarihsel Gelişimi Ve Biyolojik KavramlarDocument67 pagesTıbbi Biyolojinin Tarihsel Gelişimi Ve Biyolojik KavramlarnurefsansametNo ratings yet
- Sitoloji (Hücre Biyolojisi) : Doç. Dr. Işik Didem KaragözDocument59 pagesSitoloji (Hücre Biyolojisi) : Doç. Dr. Işik Didem KaragözFurkan TaymanNo ratings yet
- Ders 2-3 Özeti-Hücre Yapısı Ve Filogenetik AğaçDocument15 pagesDers 2-3 Özeti-Hücre Yapısı Ve Filogenetik AğaçHande Nur DüzenNo ratings yet
- Hücre Bi̇yoloji̇-1Document30 pagesHücre Bi̇yoloji̇-1Theo AndersonNo ratings yet
- Hücre BiyolojisiDocument31 pagesHücre BiyolojisizuhalNo ratings yet
- 1-TIBBİ BİYOLOJİ VE GENETİK-Hücre Kavramı Ve Hücrenin BileşenleriDocument67 pages1-TIBBİ BİYOLOJİ VE GENETİK-Hücre Kavramı Ve Hücrenin BileşenlerisevenselinayyNo ratings yet
- Hafta MikrobiyolojiDocument111 pagesHafta MikrobiyolojicihandarcanNo ratings yet
- Hücre Ve Yapısı: Doç. Dr. Harun BÖCÜKDocument32 pagesHücre Ve Yapısı: Doç. Dr. Harun BÖCÜKSanem AkdağNo ratings yet
- Deney 2Document10 pagesDeney 2ranida lcfNo ratings yet
- 1HÜCRENİN TARİHİ GELİŞİMİ, HÜCRE TEORİSİ, PROKARYOT Ve ÖKARYOT HÜCREDocument2 pages1HÜCRENİN TARİHİ GELİŞİMİ, HÜCRE TEORİSİ, PROKARYOT Ve ÖKARYOT HÜCRElt8746788No ratings yet
- Eczacilik Gi̇ri̇şDocument50 pagesEczacilik Gi̇ri̇şZzzz nnNo ratings yet
- Moleküler Biyolojiye GirişDocument25 pagesMoleküler Biyolojiye GirişKerim YılkıNo ratings yet
- Canlılar Alemi&mikroorganizmalarDocument104 pagesCanlılar Alemi&mikroorganizmalarDuygu AkyolNo ratings yet
- Prokaryot Ve Ökaryot 14.10.2016Document76 pagesProkaryot Ve Ökaryot 14.10.2016ISLAM MHANNANo ratings yet
- Hafta2 Bakteri-Yapıları1Document71 pagesHafta2 Bakteri-Yapıları1emirhanNo ratings yet
- Hücrenin Temel Yapısal ÖzellikleriDocument27 pagesHücrenin Temel Yapısal Özelliklerimehmet yiğit kıranNo ratings yet
- 1 - Tarihçe, Giriş, Mikrop DünyasıDocument92 pages1 - Tarihçe, Giriş, Mikrop Dünyasıaleynayk351No ratings yet
- 5 Dbdci 4 y 2 CyDocument16 pages5 Dbdci 4 y 2 CyEnis KartalNo ratings yet
- Hücre 3Document11 pagesHücre 3kotanbuket3No ratings yet
- Hücre Teorisi Prokaryotlar Ve ÖkaryotlarDocument16 pagesHücre Teorisi Prokaryotlar Ve ÖkaryotlarZeynep AktaşNo ratings yet
- OBRL'de Biyon-Biyogenez Üzerine Araştırmalar Ve Seminerler: İlerleme Raporu, James DeMeoDocument17 pagesOBRL'de Biyon-Biyogenez Üzerine Araştırmalar Ve Seminerler: İlerleme Raporu, James DeMeoSerdar Çetin50% (2)
- HücreDocument4 pagesHücrelloplNo ratings yet
- 4 HücreDocument139 pages4 Hücrehayripotir99No ratings yet
- 2) İlk Hücreni̇n Oluşumu, Prokaryoti̇k HücrelerDocument87 pages2) İlk Hücreni̇n Oluşumu, Prokaryoti̇k HücrelerRufat GozalovNo ratings yet
- Bilimsel Çalışma Ve Canlıların Ortak ÖzellikleriDocument32 pagesBilimsel Çalışma Ve Canlıların Ortak ÖzellikleriÖMER YANIKNo ratings yet
- Genel Biyoloji Lab 3. HaftaDocument41 pagesGenel Biyoloji Lab 3. Haftamucahitc259No ratings yet
- BelgeDocument4 pagesBelgekk6630199No ratings yet
- Çıkarılacak Biyokimya Hücre Ve Biyolojik Zarlar 2022Document71 pagesÇıkarılacak Biyokimya Hücre Ve Biyolojik Zarlar 2022Ahmet FirinNo ratings yet
- Hucre-1 Hucre Zary Sitoplazma Ve Organeller-DonuyturulduDocument8 pagesHucre-1 Hucre Zary Sitoplazma Ve Organeller-Donuyturulduapi-644562249No ratings yet
- Histolojiye GirişDocument33 pagesHistolojiye GirişSareNo ratings yet
- Prokaryot, Ökaryot Ve Virüs Dr. Behice ZERENDocument44 pagesProkaryot, Ökaryot Ve Virüs Dr. Behice ZERENbehice zerenNo ratings yet
- BMM 352 Ders 1Document34 pagesBMM 352 Ders 1melisakd55No ratings yet
- Adf 5Document8 pagesAdf 5krtpkrdNo ratings yet
- 1.deney HücreDocument57 pages1.deney HücreDIDAR YAZHANOVNo ratings yet
- 2hafta Hucre Ve SiniflandirmaDocument35 pages2hafta Hucre Ve SiniflandirmaSelim sarıgülNo ratings yet
- 9.sınıf8.not Tungsten DayiDocument4 pages9.sınıf8.not Tungsten DayiBETÜL KurtoğluNo ratings yet
- 2020.11.25 11.00 Bakterilerin Morfolojisi, Bakteri Hücresinin Yapısı, Sitoplazma, Sitoplazma Zarı 38Document4 pages2020.11.25 11.00 Bakterilerin Morfolojisi, Bakteri Hücresinin Yapısı, Sitoplazma, Sitoplazma Zarı 38A AaNo ratings yet
- Canlilarin Ortak Özelk.Document6 pagesCanlilarin Ortak Özelk.Reika -No ratings yet
- Hafta - Hücre OrganelleriDocument4 pagesHafta - Hücre Organelleriburhan CeNo ratings yet
- Ders 2Document49 pagesDers 2Hooman SavehdarbandsariNo ratings yet
- Ders3 - Canliligin Temel Birimi Hucreler Ve Cesitleri Ve Kendi Ekosistemleri Ile Olan IliskileriDocument13 pagesDers3 - Canliligin Temel Birimi Hucreler Ve Cesitleri Ve Kendi Ekosistemleri Ile Olan Iliskileriocakhande2No ratings yet
- Ders 2 HÜCREDocument29 pagesDers 2 HÜCREnisatugcebNo ratings yet
- Arguman Arsivi PDFDocument194 pagesArguman Arsivi PDFasığırcı_2No ratings yet
- Nukleik Asitler AYTDocument15 pagesNukleik Asitler AYTMüslümizmNo ratings yet
- Temel Ve Klinik Bilimler: Ders Notlari 1.CİLTDocument16 pagesTemel Ve Klinik Bilimler: Ders Notlari 1.CİLTÖZKAN ÇİÇEKNo ratings yet
- Biyo 103 Lab2Document8 pagesBiyo 103 Lab2Ali VeliNo ratings yet
- HÜCRE MEMBRANI Ve MOLEKÜLER ÖZELLİKLERİDocument59 pagesHÜCRE MEMBRANI Ve MOLEKÜLER ÖZELLİKLERİmobin.kouhi1No ratings yet
- 1 Mikrobiyolojiye Giriş, Mikroorganizmaların Yapısı Ve Sınıflandırılması (Uyumluluk Modu) - 24Document58 pages1 Mikrobiyolojiye Giriş, Mikroorganizmaların Yapısı Ve Sınıflandırılması (Uyumluluk Modu) - 24Adam SandlerNo ratings yet
- Hucrenin IçiDocument34 pagesHucrenin Içizrmlseii.0No ratings yet
- Hücre 1Document136 pagesHücre 1Kaan mert AktaşNo ratings yet
- Biyokimya Ders NotlarıDocument23 pagesBiyokimya Ders Notlarısuha_sahin3590No ratings yet
- TKB 1 Sitoloji Hücre BiyolojisiDocument91 pagesTKB 1 Sitoloji Hücre BiyolojisiCan ArasNo ratings yet
- Tıbbi Mikrobiyoloji I: Patojenler ve İnsan MikrobiyomuFrom EverandTıbbi Mikrobiyoloji I: Patojenler ve İnsan MikrobiyomuRating: 5 out of 5 stars5/5 (1)
- Kaslar Hakkında Genel BilgilerDocument44 pagesKaslar Hakkında Genel BilgilerunejsaNo ratings yet
- Cikan YollarDocument47 pagesCikan YollarunejsaNo ratings yet
- Kapiller Membrandan Madde Gecisi-2020-DonusturulduDocument36 pagesKapiller Membrandan Madde Gecisi-2020-DonusturulduunejsaNo ratings yet
- Fossatemporalisfossainfratemporalisfossapterygopalatina 130722121845 Phpapp02Document47 pagesFossatemporalisfossainfratemporalisfossapterygopalatina 130722121845 Phpapp02unejsaNo ratings yet