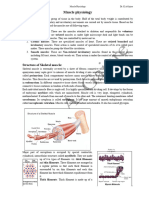
Download as pdf or txt
You might also like
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5834)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1093)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (852)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (590)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (903)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (541)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (350)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (824)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (122)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (405)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- Robbins Pathology - Chapter 1 TransDocument11 pagesRobbins Pathology - Chapter 1 Transnath nathNo ratings yet
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (74)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- Food Chemistry: Malgorzata Korzeniowska, Imelda W.Y. Cheung, Eunice C.Y. Li-ChanDocument9 pagesFood Chemistry: Malgorzata Korzeniowska, Imelda W.Y. Cheung, Eunice C.Y. Li-Chanari wawastoNo ratings yet
- PDF Seafoods Chemistry Processing Technology and Quality Fereidoon Shahidi Ebook Full ChapterDocument53 pagesPDF Seafoods Chemistry Processing Technology and Quality Fereidoon Shahidi Ebook Full Chaptersandra.millican277100% (1)
- Bio 150 Trans 3.1Document13 pagesBio 150 Trans 3.1Jolaine ValloNo ratings yet
- Physiology of Labor PDFDocument68 pagesPhysiology of Labor PDFAJ MendozaNo ratings yet
- Musculoskelet Al System - Muscles: Year 11 Human Biology ATAR Unit 1 & 2Document22 pagesMusculoskelet Al System - Muscles: Year 11 Human Biology ATAR Unit 1 & 2Anonymous vwR8xcIxNo ratings yet
- Biotech 2021Document473 pagesBiotech 2021First NameNo ratings yet
- 2Document17 pages2이어진No ratings yet
- The Cytoskeleton: Cell Biology 1st SemesterDocument22 pagesThe Cytoskeleton: Cell Biology 1st SemesterJason CenaNo ratings yet
- Perspectives: Turing's Next Steps: The Mechanochemical Basis of MorphogenesisDocument7 pagesPerspectives: Turing's Next Steps: The Mechanochemical Basis of MorphogenesisokafoieahovanNo ratings yet
- Equipo 6Document7 pagesEquipo 6Iris CsmNo ratings yet
- 11 Biology Notes ch20 Locomotion and Movement PDFDocument3 pages11 Biology Notes ch20 Locomotion and Movement PDFRamachandranPerumalNo ratings yet
- 1 MusclesDocument29 pages1 MusclesGideon CavidaNo ratings yet
- The Cytoskeleton Is A Network of Filament and Tubules That Extends Throughout A CellDocument9 pagesThe Cytoskeleton Is A Network of Filament and Tubules That Extends Throughout A CellHania GulNo ratings yet
- LSB231.5 Muscle Tissue Physiology Student Version SlidesDocument93 pagesLSB231.5 Muscle Tissue Physiology Student Version SlidesJohn GoulterNo ratings yet
- Physiology06 Muscle CellDocument28 pagesPhysiology06 Muscle CellsmpoojasubashNo ratings yet
- Cell Signaling 5 LessonDocument66 pagesCell Signaling 5 LessonPrathamesh KulkarniNo ratings yet
- CH 38 Cellular MotilityDocument19 pagesCH 38 Cellular MotilityerichaasNo ratings yet
- 589bc774 Df1d 47c6 Bc97 B5fd73030ca6muscle PhysiologyDocument3 pages589bc774 Df1d 47c6 Bc97 B5fd73030ca6muscle PhysiologyMalti SharmaNo ratings yet
- ANPH Midterm NotesDocument44 pagesANPH Midterm NotesClaudine Jo B. TalabocNo ratings yet
- Chapter 16 - The Cytoskeleton - 112612Document56 pagesChapter 16 - The Cytoskeleton - 112612Sohan rautNo ratings yet
- Exam IiiDocument22 pagesExam IiiJay ZNo ratings yet
- 3D. Muscle TissueDocument47 pages3D. Muscle Tissuejaf123xyNo ratings yet
- Chapter 20Document6 pagesChapter 20netram rajora100% (1)
- The Muscular System Anaphy LabDocument12 pagesThe Muscular System Anaphy LabHannah Grace CorveraNo ratings yet
- Locomotion and MovementDocument20 pagesLocomotion and MovementChloe AranasNo ratings yet
- Muscle Tissue SummaryDocument2 pagesMuscle Tissue SummaryLindenScholesNo ratings yet
- Explanations For Every Single Question For Every Userful Resource For The MCATDocument294 pagesExplanations For Every Single Question For Every Userful Resource For The MCATThunder BlastNo ratings yet
- Brownian Motion HandoutDocument23 pagesBrownian Motion HandoutSehrelilNo ratings yet
- Vladimir Zatsiorsky, Boris Prilutsky-Biomechanics of Skeletal Muscles-Human Kinetics (2012) PDFDocument536 pagesVladimir Zatsiorsky, Boris Prilutsky-Biomechanics of Skeletal Muscles-Human Kinetics (2012) PDFJoao CunhaNo ratings yet