
Piperlongumine Induces Apoptosis and Autophagy in Human Lung Cancer Cells Through Inhibition of PI3K/Akt/mTOR Pathway
Piperlongumine Induces Apoptosis and Autophagy in Human Lung Cancer Cells Through Inhibition of PI3K/Akt/mTOR Pathway
You might also like
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5823)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1093)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (852)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (590)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (898)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (541)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (349)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (823)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (122)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (403)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (74)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- 500 Single Best Answers in MedicineDocument443 pages500 Single Best Answers in Medicineasma .sassi100% (3)
- Glomerular DiseasesDocument211 pagesGlomerular DiseasesAdrián Beltrán67% (3)
- Copper Oxide Based Nanostructures For Improved Solar Cell EfficiencyDocument9 pagesCopper Oxide Based Nanostructures For Improved Solar Cell Efficiencyioan_vNo ratings yet
- Effect of Light Intensity - PVEducationDocument3 pagesEffect of Light Intensity - PVEducationioan_vNo ratings yet
- Nonclassical Light GenerationDocument12 pagesNonclassical Light Generationioan_vNo ratings yet
- Light Concentration Effect On PV Performance and EfficiencyDocument5 pagesLight Concentration Effect On PV Performance and Efficiencyioan_vNo ratings yet
- User Guide J2me Sony-Ericsson 2004Document134 pagesUser Guide J2me Sony-Ericsson 2004ioan_vNo ratings yet
- Physical Properties of RefrigerantsDocument2 pagesPhysical Properties of Refrigerantsioan_vNo ratings yet
- Heat of Solution DataDocument2 pagesHeat of Solution Dataioan_vNo ratings yet
- Carbon-Steam Reaction at Elevated TemperatureDocument4 pagesCarbon-Steam Reaction at Elevated Temperatureioan_vNo ratings yet
- Reduction and Oxidation PotentialDocument1 pageReduction and Oxidation Potentialioan_vNo ratings yet
- A Miller Heavy WaterDocument14 pagesA Miller Heavy Waterioan_vNo ratings yet
- 8DDocument10 pages8DaminNo ratings yet
- Section 3: Nomenclature of TumorDocument71 pagesSection 3: Nomenclature of TumorIsuri GanNo ratings yet
- Ayurveda AromatherapyDocument8 pagesAyurveda AromatherapyzbhabhaNo ratings yet
- Lung Cancer Risk Factors and EpidemiologyDocument21 pagesLung Cancer Risk Factors and EpidemiologycarolinapolotorresNo ratings yet
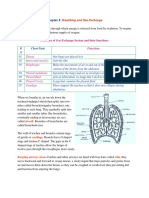
- Breathing and Gas Exchange: RespirationDocument4 pagesBreathing and Gas Exchange: RespirationAbubakar Siddiq RaminNo ratings yet
- Benign Disease of The Esophagus PDFDocument4 pagesBenign Disease of The Esophagus PDFSpecialNameNo ratings yet
- Portfolio NFDN 2004 1Document4 pagesPortfolio NFDN 2004 1api-306095474100% (1)
- Nabothian CystsDocument24 pagesNabothian CystsVirna SeptianaNo ratings yet
- 50 YEARS OF IDEAS in Health Care Buildin PDFDocument80 pages50 YEARS OF IDEAS in Health Care Buildin PDFAndrada Iulia NeagNo ratings yet
- AIIMS RECALL NOVEMBER 2019 - Updated Version 2Document83 pagesAIIMS RECALL NOVEMBER 2019 - Updated Version 2Prasad KambleNo ratings yet
- DALY Ischaemic and Rheumatic Diseases Country WiseDocument23 pagesDALY Ischaemic and Rheumatic Diseases Country Wisesagvid25No ratings yet
- Acute AppendicitisDocument6 pagesAcute AppendicitisJoel O'neil Alastair BrownNo ratings yet
- Cervical Cancer: Dr. Sushma DhakalDocument137 pagesCervical Cancer: Dr. Sushma DhakalBhattarai ShrinkhalaNo ratings yet
- EndometriosisDocument29 pagesEndometriosisJohnnyDeepNo ratings yet
- 100 Most Important DrugsDocument13 pages100 Most Important Drugsngopya djiki67% (3)
- Beat Autoimmune - Quickstart GuideDocument26 pagesBeat Autoimmune - Quickstart GuidedaveNo ratings yet
- 86 Different Kinds of Herbal TeaDocument10 pages86 Different Kinds of Herbal TeawhitecreamcoffeeNo ratings yet
- Pulmonary TuberculosisDocument2 pagesPulmonary TuberculosissoykapitanNo ratings yet
- European Journal of Obstetrics & Gynecology and Reproductive BiologyDocument5 pagesEuropean Journal of Obstetrics & Gynecology and Reproductive BiologyChennieWongNo ratings yet
- TMR of The GCR - Transformation, Modernization, Reindustrialisation of The Gauteng City-RegionDocument74 pagesTMR of The GCR - Transformation, Modernization, Reindustrialisation of The Gauteng City-RegionPatricia TheronNo ratings yet
- Child Guard Factsheet 210613Document4 pagesChild Guard Factsheet 210613nusthe27450% (1)
- 02 - Dental Focal InfectionDocument13 pages02 - Dental Focal Infectionmichal ben meronNo ratings yet
- Gastrointestinal DiseaseDocument20 pagesGastrointestinal DiseasenarenmaniamNo ratings yet
- Alternative and Complementary Medicine - Integrative Medicine and Its FutureDocument1 pageAlternative and Complementary Medicine - Integrative Medicine and Its FuturenaveenNo ratings yet
- Important Numbers For SMLEDocument3 pagesImportant Numbers For SMLEAkpevwe EmefeNo ratings yet
- TG42 Report PDFDocument68 pagesTG42 Report PDFIonio PelagosNo ratings yet
- Rundown PIT IAPIDocument2 pagesRundown PIT IAPIMade RusmanaNo ratings yet
- Draft List References Supporting Assessment Curcuma Longa L C Domestica Valeton Rhizome Revision 1 - enDocument9 pagesDraft List References Supporting Assessment Curcuma Longa L C Domestica Valeton Rhizome Revision 1 - enka waiiNo ratings yet
- Book, 3rd Spring Meeting Bali, IndonesiaDocument45 pagesBook, 3rd Spring Meeting Bali, Indonesia_wijonoNo ratings yet

















































































Download as pdf or txt
You might also like
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5823)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1093)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (852)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (590)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (898)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (541)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (349)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (823)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (122)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (403)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (74)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- 500 Single Best Answers in MedicineDocument443 pages500 Single Best Answers in Medicineasma .sassi100% (3)
- Glomerular DiseasesDocument211 pagesGlomerular DiseasesAdrián Beltrán67% (3)
- Copper Oxide Based Nanostructures For Improved Solar Cell EfficiencyDocument9 pagesCopper Oxide Based Nanostructures For Improved Solar Cell Efficiencyioan_vNo ratings yet
- Effect of Light Intensity - PVEducationDocument3 pagesEffect of Light Intensity - PVEducationioan_vNo ratings yet
- Nonclassical Light GenerationDocument12 pagesNonclassical Light Generationioan_vNo ratings yet
- Light Concentration Effect On PV Performance and EfficiencyDocument5 pagesLight Concentration Effect On PV Performance and Efficiencyioan_vNo ratings yet
- User Guide J2me Sony-Ericsson 2004Document134 pagesUser Guide J2me Sony-Ericsson 2004ioan_vNo ratings yet
- Physical Properties of RefrigerantsDocument2 pagesPhysical Properties of Refrigerantsioan_vNo ratings yet
- Heat of Solution DataDocument2 pagesHeat of Solution Dataioan_vNo ratings yet
- Carbon-Steam Reaction at Elevated TemperatureDocument4 pagesCarbon-Steam Reaction at Elevated Temperatureioan_vNo ratings yet
- Reduction and Oxidation PotentialDocument1 pageReduction and Oxidation Potentialioan_vNo ratings yet
- A Miller Heavy WaterDocument14 pagesA Miller Heavy Waterioan_vNo ratings yet
- 8DDocument10 pages8DaminNo ratings yet
- Section 3: Nomenclature of TumorDocument71 pagesSection 3: Nomenclature of TumorIsuri GanNo ratings yet
- Ayurveda AromatherapyDocument8 pagesAyurveda AromatherapyzbhabhaNo ratings yet
- Lung Cancer Risk Factors and EpidemiologyDocument21 pagesLung Cancer Risk Factors and EpidemiologycarolinapolotorresNo ratings yet
- Breathing and Gas Exchange: RespirationDocument4 pagesBreathing and Gas Exchange: RespirationAbubakar Siddiq RaminNo ratings yet
- Benign Disease of The Esophagus PDFDocument4 pagesBenign Disease of The Esophagus PDFSpecialNameNo ratings yet
- Portfolio NFDN 2004 1Document4 pagesPortfolio NFDN 2004 1api-306095474100% (1)
- Nabothian CystsDocument24 pagesNabothian CystsVirna SeptianaNo ratings yet
- 50 YEARS OF IDEAS in Health Care Buildin PDFDocument80 pages50 YEARS OF IDEAS in Health Care Buildin PDFAndrada Iulia NeagNo ratings yet
- AIIMS RECALL NOVEMBER 2019 - Updated Version 2Document83 pagesAIIMS RECALL NOVEMBER 2019 - Updated Version 2Prasad KambleNo ratings yet
- DALY Ischaemic and Rheumatic Diseases Country WiseDocument23 pagesDALY Ischaemic and Rheumatic Diseases Country Wisesagvid25No ratings yet
- Acute AppendicitisDocument6 pagesAcute AppendicitisJoel O'neil Alastair BrownNo ratings yet
- Cervical Cancer: Dr. Sushma DhakalDocument137 pagesCervical Cancer: Dr. Sushma DhakalBhattarai ShrinkhalaNo ratings yet
- EndometriosisDocument29 pagesEndometriosisJohnnyDeepNo ratings yet
- 100 Most Important DrugsDocument13 pages100 Most Important Drugsngopya djiki67% (3)
- Beat Autoimmune - Quickstart GuideDocument26 pagesBeat Autoimmune - Quickstart GuidedaveNo ratings yet
- 86 Different Kinds of Herbal TeaDocument10 pages86 Different Kinds of Herbal TeawhitecreamcoffeeNo ratings yet
- Pulmonary TuberculosisDocument2 pagesPulmonary TuberculosissoykapitanNo ratings yet
- European Journal of Obstetrics & Gynecology and Reproductive BiologyDocument5 pagesEuropean Journal of Obstetrics & Gynecology and Reproductive BiologyChennieWongNo ratings yet
- TMR of The GCR - Transformation, Modernization, Reindustrialisation of The Gauteng City-RegionDocument74 pagesTMR of The GCR - Transformation, Modernization, Reindustrialisation of The Gauteng City-RegionPatricia TheronNo ratings yet
- Child Guard Factsheet 210613Document4 pagesChild Guard Factsheet 210613nusthe27450% (1)
- 02 - Dental Focal InfectionDocument13 pages02 - Dental Focal Infectionmichal ben meronNo ratings yet
- Gastrointestinal DiseaseDocument20 pagesGastrointestinal DiseasenarenmaniamNo ratings yet
- Alternative and Complementary Medicine - Integrative Medicine and Its FutureDocument1 pageAlternative and Complementary Medicine - Integrative Medicine and Its FuturenaveenNo ratings yet
- Important Numbers For SMLEDocument3 pagesImportant Numbers For SMLEAkpevwe EmefeNo ratings yet
- TG42 Report PDFDocument68 pagesTG42 Report PDFIonio PelagosNo ratings yet
- Rundown PIT IAPIDocument2 pagesRundown PIT IAPIMade RusmanaNo ratings yet
- Draft List References Supporting Assessment Curcuma Longa L C Domestica Valeton Rhizome Revision 1 - enDocument9 pagesDraft List References Supporting Assessment Curcuma Longa L C Domestica Valeton Rhizome Revision 1 - enka waiiNo ratings yet
- Book, 3rd Spring Meeting Bali, IndonesiaDocument45 pagesBook, 3rd Spring Meeting Bali, Indonesia_wijonoNo ratings yet