
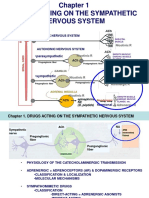
Dopamine R Gic
Dopamine R Gic
You might also like
- Anorexia Nervosa - A Multi-Disciplinary Approach - A. Mancini (Nova, 2010) WWDocument265 pagesAnorexia Nervosa - A Multi-Disciplinary Approach - A. Mancini (Nova, 2010) WWMada AndreiNo ratings yet
- Dopamine-Deficient MiceDocument13 pagesDopamine-Deficient Micemind2analysisNo ratings yet
- Dopaminergic Neurons: Shankar J. Chinta, Julie K. AndersenDocument5 pagesDopaminergic Neurons: Shankar J. Chinta, Julie K. AndersenAlejandra CastillaNo ratings yet
- The Role of RNA DNA and PROTEIN in The Transmission of TraitsDocument21 pagesThe Role of RNA DNA and PROTEIN in The Transmission of TraitsrebugiocosmeNo ratings yet
- Amine NeurotransmittersDocument7 pagesAmine NeurotransmittersFrancisco BecerraNo ratings yet
- Catecholamines: (Dopamine (DA), Norepinephrine (NE), Epinephrine (EPI) )Document30 pagesCatecholamines: (Dopamine (DA), Norepinephrine (NE), Epinephrine (EPI) )Imrana AamirNo ratings yet
- Protein SynthesisDocument67 pagesProtein SynthesisLian Rose MendozaNo ratings yet
- 3 ANSDrugsDocument75 pages3 ANSDrugsLinearNo ratings yet
- Central DogmaDocument52 pagesCentral DogmaMark Vincent Rei VillacruzNo ratings yet
- Neurotransmitter 2Document37 pagesNeurotransmitter 2Preety ShresthaNo ratings yet
- Chapter-6 Molecular Basis of InheritanceDocument10 pagesChapter-6 Molecular Basis of InheritancebpmbhamoraNo ratings yet
- Bylund 2014Document3 pagesBylund 2014Javier RiveraNo ratings yet
- 2 AdoneDocument14 pages2 AdoneJoel BotinNo ratings yet
- Genes and Chromosomes: Lehninger. Principles of Biochemistry. by Nelson and Cox, 5 Edition W.H. Freeman and CompanyDocument121 pagesGenes and Chromosomes: Lehninger. Principles of Biochemistry. by Nelson and Cox, 5 Edition W.H. Freeman and CompanyAmeylia KrisNo ratings yet
- W4-1 Transcription StuDocument25 pagesW4-1 Transcription Stu23005965No ratings yet
- Biochemistry Kaplan (AutoRecovered)Document108 pagesBiochemistry Kaplan (AutoRecovered)Khoa VõNo ratings yet
- Protein Synthesis Completed Green PacketDocument6 pagesProtein Synthesis Completed Green PacketrltradetrashNo ratings yet
- Mitochondrial Dna and DiseaseDocument16 pagesMitochondrial Dna and DiseaseNathaly LapoNo ratings yet
- Neuro TransmittersDocument7 pagesNeuro TransmittersNitishaNo ratings yet
- Genetics: Unit III - Living Things and Its Environment - Heredity: Inheritance & VariationDocument4 pagesGenetics: Unit III - Living Things and Its Environment - Heredity: Inheritance & VariationKirito TobioNo ratings yet
- Reviewer of A PsychologyDocument55 pagesReviewer of A PsychologyCAMMILLE EDZ FERRAS SELONGNo ratings yet
- 1 Sympathomimetic DrugsDocument35 pages1 Sympathomimetic DrugsFatima ZahraNo ratings yet
- Overview of Molecular Biology: GenomeDocument49 pagesOverview of Molecular Biology: GenomefachriybalafifNo ratings yet
- Pharmacology 011 - ANS 3Document9 pagesPharmacology 011 - ANS 3Kaye NeeNo ratings yet
- Neurotransmitter Transporters: Fruitful Targets For CNS Drug DiscoveryDocument6 pagesNeurotransmitter Transporters: Fruitful Targets For CNS Drug Discoveryochi rossyNo ratings yet
- DNA RNA NJLKJLJDocument5 pagesDNA RNA NJLKJLJDanekka TanNo ratings yet
- An Overview of Mammalian Mitochondrial DNA Replication Mechanisms 2018Document11 pagesAn Overview of Mammalian Mitochondrial DNA Replication Mechanisms 2018AlienNo ratings yet
- Dopamina No AprendizadoDocument12 pagesDopamina No AprendizadoGladys MariottoNo ratings yet
- Dna, Rna, Dan Protein 1Document26 pagesDna, Rna, Dan Protein 1R yNo ratings yet
- The Chemical Basis of Heredity: Yvonne Hazel M. Fortun Mscied-Biology 1Document132 pagesThe Chemical Basis of Heredity: Yvonne Hazel M. Fortun Mscied-Biology 1Yvonne Hazel FortunNo ratings yet
- Nuceic Acid ProjectDocument2 pagesNuceic Acid ProjectCaryl Alvarado SilangNo ratings yet
- Endocrine Respiratory Reproductive Excretory Urinary Nervous Immune SysytemDocument185 pagesEndocrine Respiratory Reproductive Excretory Urinary Nervous Immune SysytemEms TeopeNo ratings yet
- Biochemistry of NeurotransmittersDocument7 pagesBiochemistry of NeurotransmittersHyphophysis 2015No ratings yet
- Nucleotides, Nucleic Acids, and HeredityDocument42 pagesNucleotides, Nucleic Acids, and HeredityLyssaMarieKathryneEgeNo ratings yet
- Self Study Assignment On Marker Enzymes, Semiautonomous - PPTX 1740512Document20 pagesSelf Study Assignment On Marker Enzymes, Semiautonomous - PPTX 1740512aswinNo ratings yet
- DNA StructureDocument83 pagesDNA StructureRoliane RugaNo ratings yet
- LG 11-14Document56 pagesLG 11-14hasan.hmu2004No ratings yet
- Neurotransmitters Implicated in Psychiatry: MENTOR-Dr. Lokesh Shekhawat Sir. PRESENTOR - Dr. Vibha TomarDocument79 pagesNeurotransmitters Implicated in Psychiatry: MENTOR-Dr. Lokesh Shekhawat Sir. PRESENTOR - Dr. Vibha Tomaranon_870875350100% (1)
- Molecular Biology 01Document23 pagesMolecular Biology 01YangNo ratings yet
- Nucleic Acid Protein Synth 2022Document30 pagesNucleic Acid Protein Synth 2022Riza FirmansyahNo ratings yet
- Reviewer-in-Adv.-Biology-Nucleic-acids by JTBDocument7 pagesReviewer-in-Adv.-Biology-Nucleic-acids by JTBJoyce Fraulein T. LejosNo ratings yet
- Cells 3 PPT 2022 PDFDocument25 pagesCells 3 PPT 2022 PDFNinjaTylerFortniteBlevins 2.0No ratings yet
- Gene Expression: Replication, Transcription & Translation: Maya Dian Rakhmawatie FK Unimus - Blok 2 TA 2012/2013Document71 pagesGene Expression: Replication, Transcription & Translation: Maya Dian Rakhmawatie FK Unimus - Blok 2 TA 2012/2013Hera Vinandika PNo ratings yet
- Chromosomes: Strcture and FoldingDocument25 pagesChromosomes: Strcture and FoldingANUPAM ANAND KUMAR PANDEY MBA-INo ratings yet
- roledopamineOMCL2016 9730467Document13 pagesroledopamineOMCL2016 9730467gbautista.fuerteNo ratings yet
- The Human Genome - FinalDocument27 pagesThe Human Genome - FinalAbhi SachdevNo ratings yet
- ZilniceDocument23 pagesZilnice5cm46hysvxNo ratings yet
- HY BiochemDocument59 pagesHY BiochemLayan MohammadNo ratings yet
- Restriction Enzyme DigestionDocument3 pagesRestriction Enzyme DigestionMahathir Mohmed100% (1)
- Week11 Genbio2Document55 pagesWeek11 Genbio2PAHAM MALAPITANNo ratings yet
- vt59.2708 21321403689 - 889739862127141 - 7332965126770472921 - N.pptxdna The Human Body Recipe InfographicsDocument1 pagevt59.2708 21321403689 - 889739862127141 - 7332965126770472921 - N.pptxdna The Human Body Recipe InfographicsCassandra Bernice ReyesNo ratings yet
- Dna 1Document31 pagesDna 1Dr-Dalya ShakirNo ratings yet
- DNA and RNADocument3 pagesDNA and RNAHans PNo ratings yet
- Pharmacol Ogy PHL 313: Chapter 5-6-7 Drugs Affecting The Autonomic Nervous System (ANS)Document14 pagesPharmacol Ogy PHL 313: Chapter 5-6-7 Drugs Affecting The Autonomic Nervous System (ANS)Abdullah Al-TuraifiNo ratings yet
- TP 2 PCR 21 - 22Document50 pagesTP 2 PCR 21 - 22Vera MenezesNo ratings yet
- Lecture10 DNATranscriptionDocument40 pagesLecture10 DNATranscriptionGurlInpinkNo ratings yet
- Learner's Activity Sheet: Science (Quarter III - Week 4)Document10 pagesLearner's Activity Sheet: Science (Quarter III - Week 4)MARITESS COLLADONo ratings yet
- Nucleotides, Nucleic Acids, and Heredity: Bettelheim / Brown / Campbell / Farrell / TorresDocument55 pagesNucleotides, Nucleic Acids, and Heredity: Bettelheim / Brown / Campbell / Farrell / TorresMica BernardoNo ratings yet
- Anyone Know What This Is?: SOX-17 Transcription Factor Bound To DNA! (Note: Would Not Be As Good As The YANKS98 Factor)Document87 pagesAnyone Know What This Is?: SOX-17 Transcription Factor Bound To DNA! (Note: Would Not Be As Good As The YANKS98 Factor)Kess TurnerNo ratings yet
- CH1131 - Biomolecular Engineering - Week 2 - Transcription Translation - Aug 18 21 2015Document49 pagesCH1131 - Biomolecular Engineering - Week 2 - Transcription Translation - Aug 18 21 2015joshuaNo ratings yet
- Target Delineation Pituitary Adenoma Case Studies: DR Kanhu Charan Patro ISNOCON 2019 BhopalDocument171 pagesTarget Delineation Pituitary Adenoma Case Studies: DR Kanhu Charan Patro ISNOCON 2019 BhopalsestramitaNo ratings yet
- Maternal Thyroid Function During Pregnancy and Child Brain Morphology: A Time Window-Specific Analysis of A Prospective CohortDocument9 pagesMaternal Thyroid Function During Pregnancy and Child Brain Morphology: A Time Window-Specific Analysis of A Prospective CohortsestramitaNo ratings yet
- Immune ResponDocument30 pagesImmune ResponsestramitaNo ratings yet
- Sestramita Tuah KPJ International College of Nursing & Health SciencesDocument52 pagesSestramita Tuah KPJ International College of Nursing & Health SciencessestramitaNo ratings yet
- Renal PDFDocument16 pagesRenal PDFsestramitaNo ratings yet
- Eukaryotes: Nucleolus (The Site of Ribosome Synthesis) - in Addition To The Nucleus That Encodes GenesDocument8 pagesEukaryotes: Nucleolus (The Site of Ribosome Synthesis) - in Addition To The Nucleus That Encodes GenessestramitaNo ratings yet
- Sestramita Tuah KPJ International College of Nursing & Health SciencesDocument52 pagesSestramita Tuah KPJ International College of Nursing & Health SciencessestramitaNo ratings yet
- Endocrinology Lecture 2012Document232 pagesEndocrinology Lecture 2012Takacs CsabaNo ratings yet
- Consultation-Liaison Psychiatry (Psychosomatic Medicine) : M.Zainie Hassan A.RDocument25 pagesConsultation-Liaison Psychiatry (Psychosomatic Medicine) : M.Zainie Hassan A.RDavi DzikirianNo ratings yet
- Disorders of Water BalanceDocument64 pagesDisorders of Water BalanceRadley Jed C. PelagioNo ratings yet
- Yen Jaffes Reproductive Endocrinology E Book Physiology Pathophysiology and Clinical Management 7Th Full ChapterDocument38 pagesYen Jaffes Reproductive Endocrinology E Book Physiology Pathophysiology and Clinical Management 7Th Full Chapterjoseph.chavez825100% (27)
- Endocrine System - MedDocument124 pagesEndocrine System - Medrediet shimekachNo ratings yet
- Kumar2018 ReferenceWorkEntry EndocrineSystem PDFDocument26 pagesKumar2018 ReferenceWorkEntry EndocrineSystem PDFRamchand Bruce PhagooNo ratings yet
- Tsigos - Stress, Endocrine Physiology and Pathophysiology PDFDocument46 pagesTsigos - Stress, Endocrine Physiology and Pathophysiology PDFTrigonggoNo ratings yet
- 22nd IUNS-ICNDocument1,159 pages22nd IUNS-ICNNajwaNo ratings yet
- Neuroendocrine Control of Food IntakeDocument11 pagesNeuroendocrine Control of Food IntakeO KiNo ratings yet
- l8 Cc2 Lab Hypothalamus and Pituitary Gland Wala Nahuman PoDocument15 pagesl8 Cc2 Lab Hypothalamus and Pituitary Gland Wala Nahuman PoAlyanaNo ratings yet
- Homeostasis and Higher Brain FunctionsDocument84 pagesHomeostasis and Higher Brain Functionsbodeadumitru9261No ratings yet
- Anatomy and Function of The HypothalamusDocument13 pagesAnatomy and Function of The HypothalamusNrs Sani Sule MashiNo ratings yet
- 337 Pdfsam NMS PhysiologyDocument166 pages337 Pdfsam NMS PhysiologydrpnnreddyNo ratings yet
- Module 1: The Endocrine System: Learning ObjectivesDocument1 pageModule 1: The Endocrine System: Learning ObjectivesZeina SharkasNo ratings yet
- ENDOCRINOLOGYDocument63 pagesENDOCRINOLOGYYuni IndrianiNo ratings yet
- Neuroanatomy of Neuroendocrine SystemsDocument521 pagesNeuroanatomy of Neuroendocrine SystemsHir100% (2)
- Anatomy of The Limbic SystemDocument32 pagesAnatomy of The Limbic SystemMohammed ShaikNo ratings yet
- Hypothalamus: By: Ma. Athena C. Del Rosario-Rebulado MD September 10, 2021Document45 pagesHypothalamus: By: Ma. Athena C. Del Rosario-Rebulado MD September 10, 2021AthenaNo ratings yet
- Hypothalamus and Its Hormones, Hormones of The Pituitary GlandDocument28 pagesHypothalamus and Its Hormones, Hormones of The Pituitary GlandAnirudh AcharyaNo ratings yet
- Psychology Themes and Variations Canadian 4th Edition Weiten Test BankDocument39 pagesPsychology Themes and Variations Canadian 4th Edition Weiten Test Bankleowright2n3k100% (14)
- Chapter 20 SummaryDocument8 pagesChapter 20 SummaryNoreen B. BañagadoNo ratings yet
- DiencephalonDocument24 pagesDiencephalonHostiuc Alex RaulNo ratings yet
- Dibner Et Al 2010Document36 pagesDibner Et Al 2010SAMARAH SANTOSNo ratings yet
- Full Ebook of Pediatric Endocrinology A Clinical Handbook Dennis M Styne Online PDF All ChapterDocument69 pagesFull Ebook of Pediatric Endocrinology A Clinical Handbook Dennis M Styne Online PDF All Chaptergilliganczbch100% (5)
- Posterior Pituitary: DR Axelle Saverettiar Mbbs Physiology Department SSR Medical CollegeDocument60 pagesPosterior Pituitary: DR Axelle Saverettiar Mbbs Physiology Department SSR Medical CollegeÑäd ÉèmNo ratings yet
- The Limbic System: Juan Enrique Toro Perez UWO Fellow Neurology Residents Neurosciences Review Tutor: Jorge BurneoDocument54 pagesThe Limbic System: Juan Enrique Toro Perez UWO Fellow Neurology Residents Neurosciences Review Tutor: Jorge BurneoJuan Enrique Toro PerezNo ratings yet
- Chapter 5 - The Posterior Pituitary (Neurohypophysis)Document23 pagesChapter 5 - The Posterior Pituitary (Neurohypophysis)張宏軒No ratings yet
- Hormon Hipotalamus Dan Hipofisis Compatibility ModeDocument35 pagesHormon Hipotalamus Dan Hipofisis Compatibility ModeVennia Riskia TristiantiNo ratings yet
- Posterior Pituitary Gland Presentation - paraDocument40 pagesPosterior Pituitary Gland Presentation - parahamidNo ratings yet






























































































Download as pdf or txt
You might also like
- Anorexia Nervosa - A Multi-Disciplinary Approach - A. Mancini (Nova, 2010) WWDocument265 pagesAnorexia Nervosa - A Multi-Disciplinary Approach - A. Mancini (Nova, 2010) WWMada AndreiNo ratings yet
- Dopamine-Deficient MiceDocument13 pagesDopamine-Deficient Micemind2analysisNo ratings yet
- Dopaminergic Neurons: Shankar J. Chinta, Julie K. AndersenDocument5 pagesDopaminergic Neurons: Shankar J. Chinta, Julie K. AndersenAlejandra CastillaNo ratings yet
- The Role of RNA DNA and PROTEIN in The Transmission of TraitsDocument21 pagesThe Role of RNA DNA and PROTEIN in The Transmission of TraitsrebugiocosmeNo ratings yet
- Amine NeurotransmittersDocument7 pagesAmine NeurotransmittersFrancisco BecerraNo ratings yet
- Catecholamines: (Dopamine (DA), Norepinephrine (NE), Epinephrine (EPI) )Document30 pagesCatecholamines: (Dopamine (DA), Norepinephrine (NE), Epinephrine (EPI) )Imrana AamirNo ratings yet
- Protein SynthesisDocument67 pagesProtein SynthesisLian Rose MendozaNo ratings yet
- 3 ANSDrugsDocument75 pages3 ANSDrugsLinearNo ratings yet
- Central DogmaDocument52 pagesCentral DogmaMark Vincent Rei VillacruzNo ratings yet
- Neurotransmitter 2Document37 pagesNeurotransmitter 2Preety ShresthaNo ratings yet
- Chapter-6 Molecular Basis of InheritanceDocument10 pagesChapter-6 Molecular Basis of InheritancebpmbhamoraNo ratings yet
- Bylund 2014Document3 pagesBylund 2014Javier RiveraNo ratings yet
- 2 AdoneDocument14 pages2 AdoneJoel BotinNo ratings yet
- Genes and Chromosomes: Lehninger. Principles of Biochemistry. by Nelson and Cox, 5 Edition W.H. Freeman and CompanyDocument121 pagesGenes and Chromosomes: Lehninger. Principles of Biochemistry. by Nelson and Cox, 5 Edition W.H. Freeman and CompanyAmeylia KrisNo ratings yet
- W4-1 Transcription StuDocument25 pagesW4-1 Transcription Stu23005965No ratings yet
- Biochemistry Kaplan (AutoRecovered)Document108 pagesBiochemistry Kaplan (AutoRecovered)Khoa VõNo ratings yet
- Protein Synthesis Completed Green PacketDocument6 pagesProtein Synthesis Completed Green PacketrltradetrashNo ratings yet
- Mitochondrial Dna and DiseaseDocument16 pagesMitochondrial Dna and DiseaseNathaly LapoNo ratings yet
- Neuro TransmittersDocument7 pagesNeuro TransmittersNitishaNo ratings yet
- Genetics: Unit III - Living Things and Its Environment - Heredity: Inheritance & VariationDocument4 pagesGenetics: Unit III - Living Things and Its Environment - Heredity: Inheritance & VariationKirito TobioNo ratings yet
- Reviewer of A PsychologyDocument55 pagesReviewer of A PsychologyCAMMILLE EDZ FERRAS SELONGNo ratings yet
- 1 Sympathomimetic DrugsDocument35 pages1 Sympathomimetic DrugsFatima ZahraNo ratings yet
- Overview of Molecular Biology: GenomeDocument49 pagesOverview of Molecular Biology: GenomefachriybalafifNo ratings yet
- Pharmacology 011 - ANS 3Document9 pagesPharmacology 011 - ANS 3Kaye NeeNo ratings yet
- Neurotransmitter Transporters: Fruitful Targets For CNS Drug DiscoveryDocument6 pagesNeurotransmitter Transporters: Fruitful Targets For CNS Drug Discoveryochi rossyNo ratings yet
- DNA RNA NJLKJLJDocument5 pagesDNA RNA NJLKJLJDanekka TanNo ratings yet
- An Overview of Mammalian Mitochondrial DNA Replication Mechanisms 2018Document11 pagesAn Overview of Mammalian Mitochondrial DNA Replication Mechanisms 2018AlienNo ratings yet
- Dopamina No AprendizadoDocument12 pagesDopamina No AprendizadoGladys MariottoNo ratings yet
- Dna, Rna, Dan Protein 1Document26 pagesDna, Rna, Dan Protein 1R yNo ratings yet
- The Chemical Basis of Heredity: Yvonne Hazel M. Fortun Mscied-Biology 1Document132 pagesThe Chemical Basis of Heredity: Yvonne Hazel M. Fortun Mscied-Biology 1Yvonne Hazel FortunNo ratings yet
- Nuceic Acid ProjectDocument2 pagesNuceic Acid ProjectCaryl Alvarado SilangNo ratings yet
- Endocrine Respiratory Reproductive Excretory Urinary Nervous Immune SysytemDocument185 pagesEndocrine Respiratory Reproductive Excretory Urinary Nervous Immune SysytemEms TeopeNo ratings yet
- Biochemistry of NeurotransmittersDocument7 pagesBiochemistry of NeurotransmittersHyphophysis 2015No ratings yet
- Nucleotides, Nucleic Acids, and HeredityDocument42 pagesNucleotides, Nucleic Acids, and HeredityLyssaMarieKathryneEgeNo ratings yet
- Self Study Assignment On Marker Enzymes, Semiautonomous - PPTX 1740512Document20 pagesSelf Study Assignment On Marker Enzymes, Semiautonomous - PPTX 1740512aswinNo ratings yet
- DNA StructureDocument83 pagesDNA StructureRoliane RugaNo ratings yet
- LG 11-14Document56 pagesLG 11-14hasan.hmu2004No ratings yet
- Neurotransmitters Implicated in Psychiatry: MENTOR-Dr. Lokesh Shekhawat Sir. PRESENTOR - Dr. Vibha TomarDocument79 pagesNeurotransmitters Implicated in Psychiatry: MENTOR-Dr. Lokesh Shekhawat Sir. PRESENTOR - Dr. Vibha Tomaranon_870875350100% (1)
- Molecular Biology 01Document23 pagesMolecular Biology 01YangNo ratings yet
- Nucleic Acid Protein Synth 2022Document30 pagesNucleic Acid Protein Synth 2022Riza FirmansyahNo ratings yet
- Reviewer-in-Adv.-Biology-Nucleic-acids by JTBDocument7 pagesReviewer-in-Adv.-Biology-Nucleic-acids by JTBJoyce Fraulein T. LejosNo ratings yet
- Cells 3 PPT 2022 PDFDocument25 pagesCells 3 PPT 2022 PDFNinjaTylerFortniteBlevins 2.0No ratings yet
- Gene Expression: Replication, Transcription & Translation: Maya Dian Rakhmawatie FK Unimus - Blok 2 TA 2012/2013Document71 pagesGene Expression: Replication, Transcription & Translation: Maya Dian Rakhmawatie FK Unimus - Blok 2 TA 2012/2013Hera Vinandika PNo ratings yet
- Chromosomes: Strcture and FoldingDocument25 pagesChromosomes: Strcture and FoldingANUPAM ANAND KUMAR PANDEY MBA-INo ratings yet
- roledopamineOMCL2016 9730467Document13 pagesroledopamineOMCL2016 9730467gbautista.fuerteNo ratings yet
- The Human Genome - FinalDocument27 pagesThe Human Genome - FinalAbhi SachdevNo ratings yet
- ZilniceDocument23 pagesZilnice5cm46hysvxNo ratings yet
- HY BiochemDocument59 pagesHY BiochemLayan MohammadNo ratings yet
- Restriction Enzyme DigestionDocument3 pagesRestriction Enzyme DigestionMahathir Mohmed100% (1)
- Week11 Genbio2Document55 pagesWeek11 Genbio2PAHAM MALAPITANNo ratings yet
- vt59.2708 21321403689 - 889739862127141 - 7332965126770472921 - N.pptxdna The Human Body Recipe InfographicsDocument1 pagevt59.2708 21321403689 - 889739862127141 - 7332965126770472921 - N.pptxdna The Human Body Recipe InfographicsCassandra Bernice ReyesNo ratings yet
- Dna 1Document31 pagesDna 1Dr-Dalya ShakirNo ratings yet
- DNA and RNADocument3 pagesDNA and RNAHans PNo ratings yet
- Pharmacol Ogy PHL 313: Chapter 5-6-7 Drugs Affecting The Autonomic Nervous System (ANS)Document14 pagesPharmacol Ogy PHL 313: Chapter 5-6-7 Drugs Affecting The Autonomic Nervous System (ANS)Abdullah Al-TuraifiNo ratings yet
- TP 2 PCR 21 - 22Document50 pagesTP 2 PCR 21 - 22Vera MenezesNo ratings yet
- Lecture10 DNATranscriptionDocument40 pagesLecture10 DNATranscriptionGurlInpinkNo ratings yet
- Learner's Activity Sheet: Science (Quarter III - Week 4)Document10 pagesLearner's Activity Sheet: Science (Quarter III - Week 4)MARITESS COLLADONo ratings yet
- Nucleotides, Nucleic Acids, and Heredity: Bettelheim / Brown / Campbell / Farrell / TorresDocument55 pagesNucleotides, Nucleic Acids, and Heredity: Bettelheim / Brown / Campbell / Farrell / TorresMica BernardoNo ratings yet
- Anyone Know What This Is?: SOX-17 Transcription Factor Bound To DNA! (Note: Would Not Be As Good As The YANKS98 Factor)Document87 pagesAnyone Know What This Is?: SOX-17 Transcription Factor Bound To DNA! (Note: Would Not Be As Good As The YANKS98 Factor)Kess TurnerNo ratings yet
- CH1131 - Biomolecular Engineering - Week 2 - Transcription Translation - Aug 18 21 2015Document49 pagesCH1131 - Biomolecular Engineering - Week 2 - Transcription Translation - Aug 18 21 2015joshuaNo ratings yet
- Target Delineation Pituitary Adenoma Case Studies: DR Kanhu Charan Patro ISNOCON 2019 BhopalDocument171 pagesTarget Delineation Pituitary Adenoma Case Studies: DR Kanhu Charan Patro ISNOCON 2019 BhopalsestramitaNo ratings yet
- Maternal Thyroid Function During Pregnancy and Child Brain Morphology: A Time Window-Specific Analysis of A Prospective CohortDocument9 pagesMaternal Thyroid Function During Pregnancy and Child Brain Morphology: A Time Window-Specific Analysis of A Prospective CohortsestramitaNo ratings yet
- Immune ResponDocument30 pagesImmune ResponsestramitaNo ratings yet
- Sestramita Tuah KPJ International College of Nursing & Health SciencesDocument52 pagesSestramita Tuah KPJ International College of Nursing & Health SciencessestramitaNo ratings yet
- Renal PDFDocument16 pagesRenal PDFsestramitaNo ratings yet
- Eukaryotes: Nucleolus (The Site of Ribosome Synthesis) - in Addition To The Nucleus That Encodes GenesDocument8 pagesEukaryotes: Nucleolus (The Site of Ribosome Synthesis) - in Addition To The Nucleus That Encodes GenessestramitaNo ratings yet
- Sestramita Tuah KPJ International College of Nursing & Health SciencesDocument52 pagesSestramita Tuah KPJ International College of Nursing & Health SciencessestramitaNo ratings yet
- Endocrinology Lecture 2012Document232 pagesEndocrinology Lecture 2012Takacs CsabaNo ratings yet
- Consultation-Liaison Psychiatry (Psychosomatic Medicine) : M.Zainie Hassan A.RDocument25 pagesConsultation-Liaison Psychiatry (Psychosomatic Medicine) : M.Zainie Hassan A.RDavi DzikirianNo ratings yet
- Disorders of Water BalanceDocument64 pagesDisorders of Water BalanceRadley Jed C. PelagioNo ratings yet
- Yen Jaffes Reproductive Endocrinology E Book Physiology Pathophysiology and Clinical Management 7Th Full ChapterDocument38 pagesYen Jaffes Reproductive Endocrinology E Book Physiology Pathophysiology and Clinical Management 7Th Full Chapterjoseph.chavez825100% (27)
- Endocrine System - MedDocument124 pagesEndocrine System - Medrediet shimekachNo ratings yet
- Kumar2018 ReferenceWorkEntry EndocrineSystem PDFDocument26 pagesKumar2018 ReferenceWorkEntry EndocrineSystem PDFRamchand Bruce PhagooNo ratings yet
- Tsigos - Stress, Endocrine Physiology and Pathophysiology PDFDocument46 pagesTsigos - Stress, Endocrine Physiology and Pathophysiology PDFTrigonggoNo ratings yet
- 22nd IUNS-ICNDocument1,159 pages22nd IUNS-ICNNajwaNo ratings yet
- Neuroendocrine Control of Food IntakeDocument11 pagesNeuroendocrine Control of Food IntakeO KiNo ratings yet
- l8 Cc2 Lab Hypothalamus and Pituitary Gland Wala Nahuman PoDocument15 pagesl8 Cc2 Lab Hypothalamus and Pituitary Gland Wala Nahuman PoAlyanaNo ratings yet
- Homeostasis and Higher Brain FunctionsDocument84 pagesHomeostasis and Higher Brain Functionsbodeadumitru9261No ratings yet
- Anatomy and Function of The HypothalamusDocument13 pagesAnatomy and Function of The HypothalamusNrs Sani Sule MashiNo ratings yet
- 337 Pdfsam NMS PhysiologyDocument166 pages337 Pdfsam NMS PhysiologydrpnnreddyNo ratings yet
- Module 1: The Endocrine System: Learning ObjectivesDocument1 pageModule 1: The Endocrine System: Learning ObjectivesZeina SharkasNo ratings yet
- ENDOCRINOLOGYDocument63 pagesENDOCRINOLOGYYuni IndrianiNo ratings yet
- Neuroanatomy of Neuroendocrine SystemsDocument521 pagesNeuroanatomy of Neuroendocrine SystemsHir100% (2)
- Anatomy of The Limbic SystemDocument32 pagesAnatomy of The Limbic SystemMohammed ShaikNo ratings yet
- Hypothalamus: By: Ma. Athena C. Del Rosario-Rebulado MD September 10, 2021Document45 pagesHypothalamus: By: Ma. Athena C. Del Rosario-Rebulado MD September 10, 2021AthenaNo ratings yet
- Hypothalamus and Its Hormones, Hormones of The Pituitary GlandDocument28 pagesHypothalamus and Its Hormones, Hormones of The Pituitary GlandAnirudh AcharyaNo ratings yet
- Psychology Themes and Variations Canadian 4th Edition Weiten Test BankDocument39 pagesPsychology Themes and Variations Canadian 4th Edition Weiten Test Bankleowright2n3k100% (14)
- Chapter 20 SummaryDocument8 pagesChapter 20 SummaryNoreen B. BañagadoNo ratings yet
- DiencephalonDocument24 pagesDiencephalonHostiuc Alex RaulNo ratings yet
- Dibner Et Al 2010Document36 pagesDibner Et Al 2010SAMARAH SANTOSNo ratings yet
- Full Ebook of Pediatric Endocrinology A Clinical Handbook Dennis M Styne Online PDF All ChapterDocument69 pagesFull Ebook of Pediatric Endocrinology A Clinical Handbook Dennis M Styne Online PDF All Chaptergilliganczbch100% (5)
- Posterior Pituitary: DR Axelle Saverettiar Mbbs Physiology Department SSR Medical CollegeDocument60 pagesPosterior Pituitary: DR Axelle Saverettiar Mbbs Physiology Department SSR Medical CollegeÑäd ÉèmNo ratings yet
- The Limbic System: Juan Enrique Toro Perez UWO Fellow Neurology Residents Neurosciences Review Tutor: Jorge BurneoDocument54 pagesThe Limbic System: Juan Enrique Toro Perez UWO Fellow Neurology Residents Neurosciences Review Tutor: Jorge BurneoJuan Enrique Toro PerezNo ratings yet
- Chapter 5 - The Posterior Pituitary (Neurohypophysis)Document23 pagesChapter 5 - The Posterior Pituitary (Neurohypophysis)張宏軒No ratings yet
- Hormon Hipotalamus Dan Hipofisis Compatibility ModeDocument35 pagesHormon Hipotalamus Dan Hipofisis Compatibility ModeVennia Riskia TristiantiNo ratings yet
- Posterior Pituitary Gland Presentation - paraDocument40 pagesPosterior Pituitary Gland Presentation - parahamidNo ratings yet