Download as pdf or txt
You might also like
- A Level Organic Chemistry NotesDocument68 pagesA Level Organic Chemistry NotesTeejay Makazhu100% (13)
- Cocaine For TropinoneDocument8 pagesCocaine For TropinoneJi ChemNo ratings yet
- Solvent Dyeing PDFDocument23 pagesSolvent Dyeing PDFDhrubo Adhikary100% (2)
- Lipase 3Document9 pagesLipase 3Angie Grados LopezNo ratings yet
- Glycosidase Enzymes Stabilization Through Immobilization Onto Nanoparticulated Bimodal OrganosilicasDocument1 pageGlycosidase Enzymes Stabilization Through Immobilization Onto Nanoparticulated Bimodal Organosilicasdadang71No ratings yet
- A Novel High Performance Enzyme For Starch LiquefactionDocument7 pagesA Novel High Performance Enzyme For Starch LiquefactionAbu ShamimNo ratings yet
- Kadri 2018Document11 pagesKadri 2018Aslam MuhammadNo ratings yet
- Basyaruddin 23 3 1Document7 pagesBasyaruddin 23 3 1NiegleNo ratings yet
- Arroz A PHB 2Document9 pagesArroz A PHB 2Bryan Roncal LlajarunaNo ratings yet
- ATPS2Document8 pagesATPS2Nicoly SilvaNo ratings yet
- 1 s2.0 S0306261912004266 Main.41Document6 pages1 s2.0 S0306261912004266 Main.41luuphuongNo ratings yet
- Extraction and Characterization of Lipase Enzymes From Bacillus Cereus (MS6) and Their Medical & Industrial ApplicationsDocument8 pagesExtraction and Characterization of Lipase Enzymes From Bacillus Cereus (MS6) and Their Medical & Industrial ApplicationsInternational Journal of Innovative Science and Research TechnologyNo ratings yet
- AMYLASEENZYMEDocument7 pagesAMYLASEENZYMEChitrikaNo ratings yet
- Partial Purification and Characterization of Amylase Enzyme Under Solid State Fermentation From Monascus SanguineusDocument7 pagesPartial Purification and Characterization of Amylase Enzyme Under Solid State Fermentation From Monascus Sanguineusalem010No ratings yet
- 2011 Article 160Document6 pages2011 Article 160jwalantkbhattNo ratings yet
- Mung Bean Protein Seminar 1Document11 pagesMung Bean Protein Seminar 1Sara PalermoNo ratings yet
- TRC Khu MauDocument14 pagesTRC Khu MauHƯƠNG NGUYỄN LÊ NGỌCNo ratings yet
- Cas Sales 2011Document9 pagesCas Sales 2011Camila NevesNo ratings yet
- PeptidoBioactivo EscamasTilapia 2021 P1BDocument10 pagesPeptidoBioactivo EscamasTilapia 2021 P1BDenis Huanca De La CruzNo ratings yet
- 10 5923 J Food 20140403 04 PDFDocument6 pages10 5923 J Food 20140403 04 PDFERIKA MARIE BECERRELNo ratings yet
- Zosterisessor Ophiocephalus, Raja Clavata Scorpaena Scrofa: Research ArticleDocument10 pagesZosterisessor Ophiocephalus, Raja Clavata Scorpaena Scrofa: Research ArticleLaila Nurul QodryNo ratings yet
- PectinasasDocument9 pagesPectinasascarolasbdNo ratings yet
- Gannou 2009Document9 pagesGannou 2009ThobiasNo ratings yet
- 1 s2.0 S1570023211007112 MainDocument6 pages1 s2.0 S1570023211007112 MainyasserNo ratings yet
- Sds-Page Com AzeiteDocument7 pagesSds-Page Com AzeiteInes LealNo ratings yet
- 4 - Choubane S., Khelil O. Et Cheba B. 2015, African Journal of BiotechnologyDocument6 pages4 - Choubane S., Khelil O. Et Cheba B. 2015, African Journal of BiotechnologyOmar KHELILNo ratings yet
- Paper 2021Document9 pagesPaper 2021Fazilah AriffinNo ratings yet
- 2008.simultaneous Non-Thermal Saccharification of Cassava Pulp by Multi-Enzyme Activity and Ethanolnext Term Fermentation by Candida TropicalisDocument6 pages2008.simultaneous Non-Thermal Saccharification of Cassava Pulp by Multi-Enzyme Activity and Ethanolnext Term Fermentation by Candida TropicalisOrban CsongorNo ratings yet
- Designing of New Acrylic Based Macroporous Superabsorbent Polymer Hydrogel and Its Suitability For Drug DeliveryDocument12 pagesDesigning of New Acrylic Based Macroporous Superabsorbent Polymer Hydrogel and Its Suitability For Drug DeliverykseshuNo ratings yet
- Research Article Bacillus SubtilisDocument10 pagesResearch Article Bacillus SubtilisDương Nguyễn Thùy DungNo ratings yet
- MesophilDocument4 pagesMesophilnurcincelikNo ratings yet
- Katalis PDFDocument13 pagesKatalis PDFkhansarafidaNo ratings yet
- A Novel Thiol-Dependent Serine Protease From Neocosmospora Sp. N1Document12 pagesA Novel Thiol-Dependent Serine Protease From Neocosmospora Sp. N1Fatema MatkawalaNo ratings yet
- Actividad Antioxidante de PeptidosDocument10 pagesActividad Antioxidante de PeptidosCarlos M. Guerra ANo ratings yet
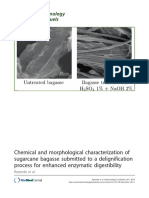
- Chemical and Morphological Characterization of Sugarcane Bagasse Submitted To A Delignification Process For Enhanced Enzymatic DigestibilityDocument19 pagesChemical and Morphological Characterization of Sugarcane Bagasse Submitted To A Delignification Process For Enhanced Enzymatic DigestibilityTabiita Huaman NorabuenaNo ratings yet
- Paper 2 RRJMB-Amylase Producing Bacteria From SalternsDocument4 pagesPaper 2 RRJMB-Amylase Producing Bacteria From SalternsSyed ShameerNo ratings yet
- Research ArticleDocument9 pagesResearch ArticleElla Gita SilvianaNo ratings yet
- En Zima Conserva Ça oDocument7 pagesEn Zima Conserva Ça oJonas TeixeiraNo ratings yet
- Si 2015Document8 pagesSi 2015ThuNo ratings yet
- Amylase Production by Bacillus Sp. Using Cassava As SubstrateDocument7 pagesAmylase Production by Bacillus Sp. Using Cassava As SubstrateMuhammad Abdur RokhimNo ratings yet
- Purification and Characterization of An Organic Solvent and Detergent-Tolerant Novel Protease Produced by Bacillus Sp. RKY3Document8 pagesPurification and Characterization of An Organic Solvent and Detergent-Tolerant Novel Protease Produced by Bacillus Sp. RKY3سید حسین عارفیNo ratings yet
- Applied Energy: J.R. Monte, M. Brienzo, A.M.F. MilagresDocument6 pagesApplied Energy: J.R. Monte, M. Brienzo, A.M.F. MilagresArif HidayatNo ratings yet
- Renewable Energy: I. Egüés, L. Serrano, D. Amendola, D.M. de Faveri, G. Spigno, J. LabidiDocument6 pagesRenewable Energy: I. Egüés, L. Serrano, D. Amendola, D.M. de Faveri, G. Spigno, J. LabidiRoshan GillNo ratings yet
- NG D NG C A Enzyme Trong Môi Trư NGDocument5 pagesNG D NG C A Enzyme Trong Môi Trư NGLuận HàNo ratings yet
- 1 s2.0 S0141813021025605 MainDocument11 pages1 s2.0 S0141813021025605 MainHanan AqoubNo ratings yet
- Binhayeeding2020 - 1Document12 pagesBinhayeeding2020 - 1Siva ForeviewNo ratings yet
- International Journal of Biological Macromolecules: S.D. Gohel, S.P. SinghDocument8 pagesInternational Journal of Biological Macromolecules: S.D. Gohel, S.P. SinghSaenz GutierrzNo ratings yet
- Yunus 2014Document10 pagesYunus 2014La Ode Muhamad RusliNo ratings yet
- Pea Protein Isolate SubstrateDocument11 pagesPea Protein Isolate SubstrateSara PalermoNo ratings yet
- Applications of Enzyme ImmobilizationDocument15 pagesApplications of Enzyme Immobilizationamrita_azam92% (13)
- Shokrkar2017 PDFDocument7 pagesShokrkar2017 PDFNissa SissariNo ratings yet
- Camargo 2014Document8 pagesCamargo 2014Julio MoralesNo ratings yet
- 1 s2.0 S135951131100448X MainDocument8 pages1 s2.0 S135951131100448X MainIngrid DantasNo ratings yet
- Research Article: Structural Analysis of Alkaline Pretreated Rice Straw For Ethanol ProductionDocument10 pagesResearch Article: Structural Analysis of Alkaline Pretreated Rice Straw For Ethanol ProductionShankar AcharNo ratings yet
- Jove Protocol 54227 Techniques For Evolution Robust Pentose Fermenting Yeast ForDocument15 pagesJove Protocol 54227 Techniques For Evolution Robust Pentose Fermenting Yeast ForAdabelle ColungaNo ratings yet
- Blue Native Electrophoresis Study On LipasesDocument2 pagesBlue Native Electrophoresis Study On LipasesRubina NeloferNo ratings yet
- Ramos-Suárez and Carreras (2014) - Use of Microalgae Residues For Biogas Production PDFDocument10 pagesRamos-Suárez and Carreras (2014) - Use of Microalgae Residues For Biogas Production PDFZouzo222No ratings yet
- Enzymatic Saccharification and Fermentation of CelDocument10 pagesEnzymatic Saccharification and Fermentation of CelSharmaNo ratings yet
- Purification and Characterization of PerDocument7 pagesPurification and Characterization of PerWilayit Ali SaediNo ratings yet
- Alkaline PhosphataseDocument7 pagesAlkaline Phosphatasemostafa sheblNo ratings yet
- Advances in Physicochemical Properties of Biopolymers: Part 2From EverandAdvances in Physicochemical Properties of Biopolymers: Part 2No ratings yet
- Practical Methods for Biocatalysis and Biotransformations 3From EverandPractical Methods for Biocatalysis and Biotransformations 3John WhittallNo ratings yet
- New Polymers for Encapsulation of Nutraceutical CompoundsFrom EverandNew Polymers for Encapsulation of Nutraceutical CompoundsJorge Carlos Ruiz RuizNo ratings yet
- Eng Journal 2Document9 pagesEng Journal 2Isnaeni RachmawatiNo ratings yet
- Wang 2018Document35 pagesWang 2018Isnaeni RachmawatiNo ratings yet
- Platyhelminthes and MolluscaDocument7 pagesPlatyhelminthes and MolluscaIsnaeni RachmawatiNo ratings yet
- Mangrove Root: Adaptations and Ecological ImportanceDocument15 pagesMangrove Root: Adaptations and Ecological ImportanceIsnaeni RachmawatiNo ratings yet
- Osher 2016Document13 pagesOsher 2016Isnaeni RachmawatiNo ratings yet
- Comparison Between Folin-Ciocalteu and Prussian Blue Assays To Estimate The Total Phenolic Content of Juices and Teas Using 96-Well MicroplatesDocument7 pagesComparison Between Folin-Ciocalteu and Prussian Blue Assays To Estimate The Total Phenolic Content of Juices and Teas Using 96-Well MicroplatesIsnaeni RachmawatiNo ratings yet
- Bialkowska Hobrzanska1990Document7 pagesBialkowska Hobrzanska1990Isnaeni RachmawatiNo ratings yet
- Chemotaxonomy in Bacterial Systematics: F. Seyma Gokdemir, Sumer ArasDocument13 pagesChemotaxonomy in Bacterial Systematics: F. Seyma Gokdemir, Sumer ArasIsnaeni RachmawatiNo ratings yet
- Gian Guzz A 2016Document11 pagesGian Guzz A 2016Isnaeni RachmawatiNo ratings yet
- Jurnal Sismik Eng 2Document8 pagesJurnal Sismik Eng 2Isnaeni RachmawatiNo ratings yet
- Lampiran UdangDocument3 pagesLampiran UdangIsnaeni RachmawatiNo ratings yet
- Effects of Petroleum Resins On Asphaltene Aggregation and Water-In-Oil Emulsion FormationDocument19 pagesEffects of Petroleum Resins On Asphaltene Aggregation and Water-In-Oil Emulsion FormationJonathanNo ratings yet
- Elementis-Nuosperse FX 605-TDSDocument2 pagesElementis-Nuosperse FX 605-TDSAlan González IglesiasNo ratings yet
- 2010-2004 Touch Up Paint CatalogDocument23 pages2010-2004 Touch Up Paint CatalogLucas WebbNo ratings yet
- Chapter 9. Phase DiagramsDocument8 pagesChapter 9. Phase DiagramsyshokeenNo ratings yet
- 5olasa 2022Document11 pages5olasa 2022volcano netNo ratings yet
- Fan 2019Document8 pagesFan 2019Ana Luisa GrafiaNo ratings yet
- NylonDocument9 pagesNylonchristopheNo ratings yet
- English Course Assignment (Chapter 2: Page 19-34 of The English For Science Book) ComparingDocument13 pagesEnglish Course Assignment (Chapter 2: Page 19-34 of The English For Science Book) ComparingPutri JuliantiNo ratings yet
- General ScienceDocument6 pagesGeneral ScienceSalvari VitasNo ratings yet
- Dilantil PDFDocument5 pagesDilantil PDFTaciturnoait NihilistaNo ratings yet
- Chapter 7Document3 pagesChapter 7Misbah Sajid ChaudhryNo ratings yet
- Physicochemical Properties Ch1Document30 pagesPhysicochemical Properties Ch1madhu bonamNo ratings yet
- Designing Fast Oxide-Ion Conductors Based On La2Mo2O9 by P. Lacorre, Nature (2000)Document3 pagesDesigning Fast Oxide-Ion Conductors Based On La2Mo2O9 by P. Lacorre, Nature (2000)avinash patelNo ratings yet
- Kill-Builders: D Da Aiilly yDocument17 pagesKill-Builders: D Da Aiilly yChris McIverNo ratings yet
- Structure of Atom AssignmentDocument20 pagesStructure of Atom Assignmentrajesh duaNo ratings yet
- Green Preparation of Bioplastics Based On Degradation and Chemical Modification of Lignin ResidueDocument20 pagesGreen Preparation of Bioplastics Based On Degradation and Chemical Modification of Lignin ResidueAndres VargasNo ratings yet
- Gravimetric AnalysisDocument7 pagesGravimetric AnalysisBotlhe Kgotla SamNo ratings yet
- 500 ML GBL, 500 ML GBL CleanerDocument2 pages500 ML GBL, 500 ML GBL CleanerbuygblcleanersNo ratings yet
- Method of Analysis For Sodium Citrate: PharmaguidelineDocument3 pagesMethod of Analysis For Sodium Citrate: Pharmaguidelinedimiz77No ratings yet
- Immobilization of Laccase Via Adsorption Onto Bimodal Mesoporous Zr-MOFDocument11 pagesImmobilization of Laccase Via Adsorption Onto Bimodal Mesoporous Zr-MOFAnaMariaDiazLasprillaNo ratings yet
- EXP-1: Determination of Saponification Value of Oils/ Fats SampleDocument9 pagesEXP-1: Determination of Saponification Value of Oils/ Fats SampleLokesh BhoiNo ratings yet
- Lab ManualDocument18 pagesLab ManualWnwixiejwis Usisjdx8xjwiNo ratings yet
- Mass Transport Through A Proton Exchange Membrane (Nafion) in Microbial Fuel CellsDocument8 pagesMass Transport Through A Proton Exchange Membrane (Nafion) in Microbial Fuel CellsanusachuappuNo ratings yet
- Activity #4.1Document3 pagesActivity #4.1RafaelaAndrea Horan PayteNo ratings yet
- 1.introduction To BiochemistryDocument23 pages1.introduction To BiochemistryIslam SamirNo ratings yet
- The Notes On Histochemical StainsDocument125 pagesThe Notes On Histochemical StainsDr Joswin Dsa75% (4)
- Fermentation HoneyDocument9 pagesFermentation HoneyPujaNo ratings yet