Download as pdf or txt
You might also like
- Data Tables For Alien Gene AnalysisDocument3 pagesData Tables For Alien Gene Analysisapi-382372564100% (3)
- Prokaryotic DNA ReplicationDocument5 pagesProkaryotic DNA ReplicationMg H67% (6)
- Membrane BiochemistryDocument21 pagesMembrane BiochemistryAnna SafitriNo ratings yet
- Biosynthesis of CollagenDocument40 pagesBiosynthesis of CollagenNausheerNo ratings yet
- Collagen PDFDocument19 pagesCollagen PDFJaveria KhanNo ratings yet
- Collagen: The BiomaterialDocument5 pagesCollagen: The BiomaterialSandhik RoyNo ratings yet
- Protein Structures Continued Proteins of ECM Collagen of Connective TissueDocument37 pagesProtein Structures Continued Proteins of ECM Collagen of Connective TissueAytajNo ratings yet
- Biology Notes: Proteins: CollagenDocument5 pagesBiology Notes: Proteins: CollagenMuhammad AkramNo ratings yet
- RISHUDocument30 pagesRISHURishu Mittal100% (1)
- We Are Intechopen, The World'S Leading Publisher of Open Access Books Built by Scientists, For ScientistsDocument35 pagesWe Are Intechopen, The World'S Leading Publisher of Open Access Books Built by Scientists, For ScientistsCamila Flórez IdárragaNo ratings yet
- Structure and Function of Macromolecules: 2.2.1 Carbohydrates-The Energy GiversDocument12 pagesStructure and Function of Macromolecules: 2.2.1 Carbohydrates-The Energy GiversSteven BanksNo ratings yet
- 04ProAMO LicyayoDocument8 pages04ProAMO LicyayoMohamidin MamalapatNo ratings yet
- Proteins: Classification FunctionsDocument20 pagesProteins: Classification FunctionsGeo NalobNo ratings yet
- Structural of Red CellsDocument2 pagesStructural of Red CellsDrbee10No ratings yet
- Bchn213 (M-Campus) Test 5 Su 9 and 10 MemoDocument5 pagesBchn213 (M-Campus) Test 5 Su 9 and 10 Memozinhlez518No ratings yet
- Module 3 - Lecture 3 - LectureDocument8 pagesModule 3 - Lecture 3 - LectureNi CaoNo ratings yet
- Natural BiopolymersDocument65 pagesNatural BiopolymersBensen HsuNo ratings yet
- J Ijbiomac 2017 07 013Document17 pagesJ Ijbiomac 2017 07 013Isaac GoldNo ratings yet
- Biochemistry LN05-2Document12 pagesBiochemistry LN05-2Rahaf Al-muhtasebNo ratings yet
- Collagen StructureDocument33 pagesCollagen StructureSagar De'biomimicNo ratings yet
- 2.4, 7.3 Proteins Biology HLDocument37 pages2.4, 7.3 Proteins Biology HLmunir sehwai.No ratings yet
- BCCH 4Document59 pagesBCCH 4NG SIRNo ratings yet
- Protein StructureDocument17 pagesProtein Structureutpal mohanNo ratings yet
- QUITALIG Biochemistry Lecture Module 5 ActivityDocument5 pagesQUITALIG Biochemistry Lecture Module 5 ActivityAloysius QuitaligNo ratings yet
- Chapter 4 Lecture PptsDocument82 pagesChapter 4 Lecture PptsJota AlcuadradoNo ratings yet
- BiochemistryDocument130 pagesBiochemistryshiyntumNo ratings yet
- Lecture 4 PDFDocument18 pagesLecture 4 PDFNazir KhanNo ratings yet
- Amino Acids - 2 - NoDocument9 pagesAmino Acids - 2 - NoSunny Thakur17No ratings yet
- CHEM 151 (Chapter 4)Document4 pagesCHEM 151 (Chapter 4)Chantel Acevero100% (1)
- Protein Shape and Protein PurificationDocument60 pagesProtein Shape and Protein PurificationMilena De CresentNo ratings yet
- Chap. 4B The Three-Dimensional Structure of Proteins: TopicsDocument28 pagesChap. 4B The Three-Dimensional Structure of Proteins: Topicscatalina esanuNo ratings yet
- CollagenDocument79 pagesCollagenRobins Dhakal100% (4)
- Lec 9Document15 pagesLec 9FARHAN KAMALNo ratings yet
- The Three-Dimensional Structure of Proteins: Protein Structure and FunctionDocument26 pagesThe Three-Dimensional Structure of Proteins: Protein Structure and Function김경환No ratings yet
- ProteinsDocument44 pagesProteinsSusan SylvianingrumNo ratings yet
- Extra Cellular Matrix ADocument17 pagesExtra Cellular Matrix Aapi-301896945No ratings yet
- Structures and Functions of ProteinDocument54 pagesStructures and Functions of ProteinRollyRiksantoNo ratings yet
- Chapter 4-Jnu Biomembranes-1 - 複本Document48 pagesChapter 4-Jnu Biomembranes-1 - 複本Wai Kwong ChiuNo ratings yet
- Fibrous ProteinDocument16 pagesFibrous ProteinMatth N. ErejerNo ratings yet
- Biomolecules - Protein 3Document18 pagesBiomolecules - Protein 3NinjaTylerFortniteBlevins 2.0No ratings yet
- ProteinsDocument5 pagesProteinsnadjaNo ratings yet
- Materials: Collagen Based Materials in Cosmetic Applications: A ReviewDocument15 pagesMaterials: Collagen Based Materials in Cosmetic Applications: A ReviewMauricio Tarquino FloresNo ratings yet
- Hemoglobin Is The One Protein Molecule That Knows by Everyone Which It Is One of The Most Importance and Plentiful Proteins of VertebratesDocument5 pagesHemoglobin Is The One Protein Molecule That Knows by Everyone Which It Is One of The Most Importance and Plentiful Proteins of VertebratesBilah BilahNo ratings yet
- 4 ProteinsDocument18 pages4 ProteinsjaymcornelioNo ratings yet
- Protein Structure and Folding LectureDocument18 pagesProtein Structure and Folding Lecturemaryscribd241No ratings yet
- Fibrous ProteinDocument40 pagesFibrous Proteinعبدالرحمن حاتمNo ratings yet
- CollagenDocument8 pagesCollagenkaditasookdeoNo ratings yet
- Online Practice Tests, Live Classes, Tutoring, Study Guides Q&A, Premium Content and MoreDocument15 pagesOnline Practice Tests, Live Classes, Tutoring, Study Guides Q&A, Premium Content and MoreYoAmoNYCNo ratings yet
- Application of 2 D-HPLC System For Plasma Protein SeparationDocument10 pagesApplication of 2 D-HPLC System For Plasma Protein Separationtrinhmy81No ratings yet
- Myoglobin: Myoglobin (Symbol MB or MB) Is An IronDocument8 pagesMyoglobin: Myoglobin (Symbol MB or MB) Is An IronNTA UGC-NETNo ratings yet
- Ripuseminar 171217093449Document52 pagesRipuseminar 171217093449Jun NguyễnNo ratings yet
- Protein: Globular and Fibrous ProteinDocument20 pagesProtein: Globular and Fibrous ProteinAndrewNo ratings yet
- 04model of Membranes - LecturesDocument11 pages04model of Membranes - LecturesmeliszaNo ratings yet
- ProteinsDocument9 pagesProteinsJada HartNo ratings yet
- Evaluation of Erythrocytes: Normal Ery Throcy TesDocument24 pagesEvaluation of Erythrocytes: Normal Ery Throcy Tesaixacamila3No ratings yet
- Protein Structure11Document5 pagesProtein Structure11gangmasterNo ratings yet
- Chapter 6 The Three Dimensional Structure of Proteins SummDocument48 pagesChapter 6 The Three Dimensional Structure of Proteins Summtania.delafuenteNo ratings yet
- CarbohidratosDocument8 pagesCarbohidratosElena OlmedoNo ratings yet
- Loops ProteinsDocument37 pagesLoops ProteinsSanjeev KumarNo ratings yet
- Peptides and Proteins: M.Prasad Naidu MSC Medical Biochemistry, PH.DDocument30 pagesPeptides and Proteins: M.Prasad Naidu MSC Medical Biochemistry, PH.DDr. M. Prasad NaiduNo ratings yet
- Mass Transport Answer Booklet 2017 6Document54 pagesMass Transport Answer Booklet 2017 6LucyNo ratings yet
- A-level Biology Revision: Cheeky Revision ShortcutsFrom EverandA-level Biology Revision: Cheeky Revision ShortcutsRating: 5 out of 5 stars5/5 (5)
- DM No. 667, S. 2022 - Resetting of The Schedule of Portfolio Day For SY 2022-2023Document3 pagesDM No. 667, S. 2022 - Resetting of The Schedule of Portfolio Day For SY 2022-2023LeneNo ratings yet
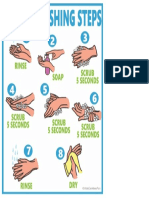
- Handwashing StepsDocument1 pageHandwashing StepsLeneNo ratings yet
- The Goose Voyage: Grade 5 Reading Comprehension WorksheetDocument4 pagesThe Goose Voyage: Grade 5 Reading Comprehension WorksheetLeneNo ratings yet
- WordsDocument1 pageWordsLeneNo ratings yet
- 5th Grade 5 Goose VoyageDocument5 pages5th Grade 5 Goose VoyageLeneNo ratings yet
- ActionDocument2 pagesActionLeneNo ratings yet
- Experiment 8 Acid-Base Titration: Pre - Lab QuestionsDocument14 pagesExperiment 8 Acid-Base Titration: Pre - Lab QuestionsLeneNo ratings yet
- SOP Preparing Sodium Hydroxide Solutions - 0Document3 pagesSOP Preparing Sodium Hydroxide Solutions - 0LeneNo ratings yet
- Report On SocDimsDocument41 pagesReport On SocDimsLeneNo ratings yet
- Mechanism of Enzyme ActionDocument10 pagesMechanism of Enzyme Actionsd2808No ratings yet
- Notes On Protein DenaturationDocument17 pagesNotes On Protein DenaturationKella OrtegaNo ratings yet
- Biochem Exam Practice QuestionsDocument12 pagesBiochem Exam Practice Questionsihack_101No ratings yet
- Price List WinSammering Pharma 2020Document1 pagePrice List WinSammering Pharma 2020shwE TVNo ratings yet
- Chart of Aminoacids Names and Abbreviations and CodonsDocument1 pageChart of Aminoacids Names and Abbreviations and Codonsstlcajun55No ratings yet
- 1 6 Monomer Drawing PracticeDocument4 pages1 6 Monomer Drawing PracticeSiddharth RajendranNo ratings yet
- Activity 5 From DNA To ProteinDocument4 pagesActivity 5 From DNA To ProteinKAREN SALLESNo ratings yet
- Himupro Protein PowderDocument1 pageHimupro Protein PowderNavi ThakurNo ratings yet
- Molecular Basis of InheritanceDocument55 pagesMolecular Basis of Inheritancearyan aggarwalNo ratings yet
- Worksheet On DNA and RNA AnswersDocument3 pagesWorksheet On DNA and RNA AnswersSelena CoronelNo ratings yet
- Chapter 2 LipidsDocument109 pagesChapter 2 LipidsTiffany Shane VallenteNo ratings yet
- DG Midterm #1 Spring 2021 From ULA ReviewDocument7 pagesDG Midterm #1 Spring 2021 From ULA ReviewRITIKA SURISETTYNo ratings yet
- Isoenzymes-Multienzyme Complex PDFDocument20 pagesIsoenzymes-Multienzyme Complex PDFintaliallitaNo ratings yet
- Gluconeogenesis AtfDocument5 pagesGluconeogenesis AtfDivyaa VisvalingamNo ratings yet
- DLL Grade 7 Video LessonDocument4 pagesDLL Grade 7 Video LessonJr CapanangNo ratings yet
- Lesson Plan Grade 8 - BiomuleculesDocument7 pagesLesson Plan Grade 8 - BiomuleculesRoxanne Reglos67% (3)
- Neptune Fish Concentrate Technical Data SheetDocument1 pageNeptune Fish Concentrate Technical Data SheetKatherine Leyla Anchayhua TorresNo ratings yet
- Insights On The Epigenetic MechanismsDocument12 pagesInsights On The Epigenetic MechanismsrafaellapordeusNo ratings yet
- Provon 395 SFL DTA - 0719-3 PDFDocument1 pageProvon 395 SFL DTA - 0719-3 PDFGomez AndreNo ratings yet
- Fatty Acids As Biocompounds: Their Role in Human Metabolism, Health and Disease - A Review. Part 1: Classification, Dietarysources and Biological FunctionsDocument14 pagesFatty Acids As Biocompounds: Their Role in Human Metabolism, Health and Disease - A Review. Part 1: Classification, Dietarysources and Biological FunctionsItan RuizNo ratings yet
- 14.respiration New BioHackDocument8 pages14.respiration New BioHackAnush KhillareNo ratings yet
- Plant Biochemistry 2 RevDocument59 pagesPlant Biochemistry 2 Revenrico andrionNo ratings yet
- Slip Test-5Document3 pagesSlip Test-5G.N. BhuvaneshwaranNo ratings yet
- Microbiology (Chapter 7: Microbial Genetics) : The Structure and Replication of GenomesDocument14 pagesMicrobiology (Chapter 7: Microbial Genetics) : The Structure and Replication of GenomesCarrie ANo ratings yet
- Harga Tayang Afifarma Non Jamkesnas 2023Document2 pagesHarga Tayang Afifarma Non Jamkesnas 2023Fadly AsriNo ratings yet
- Nucleic AcidDocument7 pagesNucleic AciddearbhupiNo ratings yet
- Ncert Line by Line Botany-Mbi Work SheetDocument17 pagesNcert Line by Line Botany-Mbi Work Sheetanusya548100% (2)