
Castanha Brazil
Castanha Brazil
You might also like
- Systems Architecture 7th Edition Burd Solutions Manual PDFDocument5 pagesSystems Architecture 7th Edition Burd Solutions Manual PDFa203025133No ratings yet
- Survival and Growth of Brazil-Nut Seedlings in Tree-Fall Gaps and Forest UnderstoryDocument10 pagesSurvival and Growth of Brazil-Nut Seedlings in Tree-Fall Gaps and Forest UnderstoryMarx Herrera MachacaNo ratings yet
- A Synopsis of The Rubiaceae of The States of Mato Grosso and Mato Grosso Do Sul, Brazil, With A Key To Genera, and A Preliminary Species ListDocument84 pagesA Synopsis of The Rubiaceae of The States of Mato Grosso and Mato Grosso Do Sul, Brazil, With A Key To Genera, and A Preliminary Species ListJúnior SenseyNo ratings yet
- Atlantic Forest To The All Americas: Biogeographical History and Divergence Times of Neotropical Ficus (Moraceae)Document51 pagesAtlantic Forest To The All Americas: Biogeographical History and Divergence Times of Neotropical Ficus (Moraceae)Francione GomesNo ratings yet
- Zuidema-Demografia de Nuez de Brazil Impactada Por El AprovechamientoDocument32 pagesZuidema-Demografia de Nuez de Brazil Impactada Por El AprovechamientoAlexis CentenoNo ratings yet
- Mares ErnstDocument20 pagesMares ErnstJoana FerreiraNo ratings yet
- Faunistic Analysis of Longhorn Beetles - Cerambycidae Coleoptera - in Cerrado and Atlantic Forest Areas Biodiversity Hotspots of BrazilDocument16 pagesFaunistic Analysis of Longhorn Beetles - Cerambycidae Coleoptera - in Cerrado and Atlantic Forest Areas Biodiversity Hotspots of BrazilMayre Ellen RodriguesNo ratings yet
- Papel de Los Mamíferos en Los Procesos de Dispersión y Depredación de Semillas de Mauritia Flexuosa (Arecaceae) en La..Document13 pagesPapel de Los Mamíferos en Los Procesos de Dispersión y Depredación de Semillas de Mauritia Flexuosa (Arecaceae) en La..Mariana Gutierrez MuneraNo ratings yet
- Lima Et Al. 2022 - NovaHedwigiaSemiaridchecklistDocument33 pagesLima Et Al. 2022 - NovaHedwigiaSemiaridchecklistJuliana LimaNo ratings yet
- Diet Specialisation in BirdsDocument17 pagesDiet Specialisation in BirdsИванАнато́лийФранкоNo ratings yet
- El Ecosistema de Dehesa Como ReservorioDocument13 pagesEl Ecosistema de Dehesa Como ReservorioIjoseangel BandinNo ratings yet
- Resturacion de IncendiosDocument9 pagesResturacion de IncendiosSalem Paco San MiguelNo ratings yet
- Biogeographic Patterns and Conservation in The South American Cerrado: A Tropical Savanna HotspotDocument12 pagesBiogeographic Patterns and Conservation in The South American Cerrado: A Tropical Savanna HotspotANDRES RICARDO LOPEZ PRECIADONo ratings yet
- A Comparison of The Wood Anatomy of 11 Species From Two Cerrado Habitats (Cerrado S.S. and Adjacent Gallery Forest) - Sonsin Et AlDocument20 pagesA Comparison of The Wood Anatomy of 11 Species From Two Cerrado Habitats (Cerrado S.S. and Adjacent Gallery Forest) - Sonsin Et Allihelih1No ratings yet
- ACFrOgDTOiHKN1mFw y7bSdQSLnMblq24oT1CulR6ZiJKiPERJ4g - CyHiovNdl AuNLd pXlWYtTXUny ZjmFgLdMJ51j2hUnRG3mCV0n3 - w4yZHMffhN7ngjTh - d08Ezh8Mv3SNY4GHwDwIbiKsDocument8 pagesACFrOgDTOiHKN1mFw y7bSdQSLnMblq24oT1CulR6ZiJKiPERJ4g - CyHiovNdl AuNLd pXlWYtTXUny ZjmFgLdMJ51j2hUnRG3mCV0n3 - w4yZHMffhN7ngjTh - d08Ezh8Mv3SNY4GHwDwIbiKsLaura MirandaNo ratings yet
- Providing Habitat For Native Mammals Through Understory Enhancement in Forestry PlantationsDocument5 pagesProviding Habitat For Native Mammals Through Understory Enhancement in Forestry Plantationsvalentina echeverriaNo ratings yet
- Janisovaal 2016 BullIAVSDocument17 pagesJanisovaal 2016 BullIAVSrangelbioNo ratings yet
- Goulart 2009Document9 pagesGoulart 2009Joana FerreiraNo ratings yet
- Non-Continuous Reproductive Phenology of Animal-Dispersed Species in Young Forest Restoration PlantingsDocument10 pagesNon-Continuous Reproductive Phenology of Animal-Dispersed Species in Young Forest Restoration PlantingsLeonardo UrruthNo ratings yet
- Bromelicolous Habit of Three Species of Rhinella Fitzinger, 1826 (Anura: Bufonidae) in Northeastern Brazil's Atlantic ForestDocument4 pagesBromelicolous Habit of Three Species of Rhinella Fitzinger, 1826 (Anura: Bufonidae) in Northeastern Brazil's Atlantic Forestvanessa e bruno becaciciNo ratings yet
- Leite Et Al 2013 - Foraging Behavior of Kaempfer's WoodpeckerDocument10 pagesLeite Et Al 2013 - Foraging Behavior of Kaempfer's WoodpeckerDianes Gomes MarcelinoNo ratings yet
- GiraldoValle CativoDocument20 pagesGiraldoValle CativoClaudia ParadaNo ratings yet
- Carlos A. Peres Luis C. Schiesari Claudia L. Dias-LemeDocument12 pagesCarlos A. Peres Luis C. Schiesari Claudia L. Dias-LemeItalo Treviño ZevallosNo ratings yet
- Galitti & Lovejoy - 2001 - The Future Atlantic Forest and Impact of LoggingDocument2 pagesGalitti & Lovejoy - 2001 - The Future Atlantic Forest and Impact of LoggingAllane SantosNo ratings yet
- Missouri Botanical Garden PressDocument34 pagesMissouri Botanical Garden PressAriadne Cristina De AntonioNo ratings yet
- The Brazilian Cerrado Vegetation and Threats To Its BiodiversityDocument8 pagesThe Brazilian Cerrado Vegetation and Threats To Its BiodiversityJackson RodriguesNo ratings yet
- Alvez-Valles Et Al. 2018 - Riqueza de Palmeiras Na AmazôniaDocument14 pagesAlvez-Valles Et Al. 2018 - Riqueza de Palmeiras Na AmazôniaGabriele FurtadoNo ratings yet
- Ecología Poblacional Del Ratón Peromyscus MexicanusDocument14 pagesEcología Poblacional Del Ratón Peromyscus MexicanusJuanNo ratings yet
- Plecevo Article 95601 en 1Document18 pagesPlecevo Article 95601 en 1Maurício GonçalvesNo ratings yet
- 1 PBDocument7 pages1 PBMylena NecoNo ratings yet
- 2 Macro PDFDocument22 pages2 Macro PDFLINA MARIA RODRIGUEZ RUIZNo ratings yet
- New Records of Macrofungi (Basidiomycota) For Colombia From A Tropical Dry ForestDocument22 pagesNew Records of Macrofungi (Basidiomycota) For Colombia From A Tropical Dry ForestLINA MARIA RODRIGUEZ RUIZNo ratings yet
- Caminha-Paiva Et Al 2022 - Fitossoc Canga QuartzitoDocument9 pagesCaminha-Paiva Et Al 2022 - Fitossoc Canga QuartzitoGeorgia SilvaNo ratings yet
- 2006-Viani, R.A.G. Et Al - Transference of Seedlings and Aloctone Young Individuals As Ecological Restoration MethodologyDocument26 pages2006-Viani, R.A.G. Et Al - Transference of Seedlings and Aloctone Young Individuals As Ecological Restoration MethodologyAdriano ItoNo ratings yet
- Linking Tropical Biology With Human DimensionsDocument5 pagesLinking Tropical Biology With Human Dimensionspabloribah7314No ratings yet
- 18157-Article Text-55730-1-10-20160202Document12 pages18157-Article Text-55730-1-10-20160202Lucìa SolerNo ratings yet
- What Do We Know About Cassis Tuberosa (Mollusca: Cassidae), A Heavily Exploited Marine Gastropod?Document14 pagesWhat Do We Know About Cassis Tuberosa (Mollusca: Cassidae), A Heavily Exploited Marine Gastropod?Murti HanafiNo ratings yet
- Artibeus Reservas Energéticas BHDocument8 pagesArtibeus Reservas Energéticas BHÉrica MunhozNo ratings yet
- Small Mammal Assemblages in A Disturbed Tropical LDocument10 pagesSmall Mammal Assemblages in A Disturbed Tropical LHeiner SaldañaNo ratings yet
- The Evolving Role of Bertholletia Excelsa in Amazonia: Contributing To Local Livelihoods and Forest ConservationDocument21 pagesThe Evolving Role of Bertholletia Excelsa in Amazonia: Contributing To Local Livelihoods and Forest ConservationMarx Herrera MachacaNo ratings yet
- Epifitas Da Pedra AzulDocument13 pagesEpifitas Da Pedra AzulAndre AssisNo ratings yet
- Edat Oliveres MilenariesDocument4 pagesEdat Oliveres MilenarieseltruferoNo ratings yet
- Fauna DarlynDocument14 pagesFauna DarlynDARLYN ASNIT LOPEZ JIMENEZNo ratings yet
- Machado 2021 Biogeography MoraceaeDocument13 pagesMachado 2021 Biogeography MoraceaeJéssica NayaraNo ratings yet
- Sinopsis Taxonómica, Ecológica y Etnobotánica Del Género Tachigali Aubl. (Leguminosae) en La Región Del Cusco, PerúDocument26 pagesSinopsis Taxonómica, Ecológica y Etnobotánica Del Género Tachigali Aubl. (Leguminosae) en La Región Del Cusco, PerúAnonymous my6XAlNo ratings yet
- Community Structure and Abundance of Small Rodents at The Wave Front of Agroforestry and Forest in Alto Beni, Bolivia - 2020Document10 pagesCommunity Structure and Abundance of Small Rodents at The Wave Front of Agroforestry and Forest in Alto Beni, Bolivia - 2020Abel Tome CaetanoNo ratings yet
- 1 s2.0 S0367253021001171 MainDocument12 pages1 s2.0 S0367253021001171 MainValeria CasiqueNo ratings yet
- Christianini 2018 Birdsfightforasinglecavity Rev Bras OrnitholDocument4 pagesChristianini 2018 Birdsfightforasinglecavity Rev Bras OrnitholjuanNo ratings yet
- Alcantara Etal 2018 Velloziaceae DiversificationDocument16 pagesAlcantara Etal 2018 Velloziaceae Diversificationdrinkto remenberNo ratings yet
- Forest Ecology and Management: A B A A C CDocument6 pagesForest Ecology and Management: A B A A C CJanaine RochaNo ratings yet
- Diversity and Seasonality of Scarabaeinae (Coleoptera Scarabaeidae) in Forest Fragments in Santa Maria, Rio Grande Do Sul, BrazilDocument19 pagesDiversity and Seasonality of Scarabaeinae (Coleoptera Scarabaeidae) in Forest Fragments in Santa Maria, Rio Grande Do Sul, BrazilEduardo ValentimNo ratings yet
- DownloadDocument9 pagesDownloadpaolospadera72No ratings yet
- Martins Et Al 2017 - Técnicas de Manejo de MelinisDocument9 pagesMartins Et Al 2017 - Técnicas de Manejo de MelinisHenrique SantosNo ratings yet
- Mammalia Firstrecordof Platyrrhinusfusciventris Chiroptera Phyllostomidaeforthe CaatingabiomeDocument6 pagesMammalia Firstrecordof Platyrrhinusfusciventris Chiroptera Phyllostomidaeforthe CaatingabiomeSara GNo ratings yet
- Diversidad Alfa y Beta de La Comunidad de Reptiles en El Complejo Cenagoso de Zapatosa, ColombiaDocument34 pagesDiversidad Alfa y Beta de La Comunidad de Reptiles en El Complejo Cenagoso de Zapatosa, ColombiaRaúl Carrillo CostaNo ratings yet
- Amburana CearensisDocument7 pagesAmburana CearensisAnthony GuerraNo ratings yet
- Tropical Rainforest Research PaperDocument5 pagesTropical Rainforest Research Papergz8qs4dn100% (1)
- 10.1515 - Mammalia 2023 0040Document7 pages10.1515 - Mammalia 2023 0040Hernani OliveiraNo ratings yet
- Ancient Transformation, Current Conservation: Traditional Forest Management On The Iriri River, Brazilian AmazoniaDocument15 pagesAncient Transformation, Current Conservation: Traditional Forest Management On The Iriri River, Brazilian AmazoniaMauricio TorresNo ratings yet
- Food Availability, Plant Diversity, and Vegetation Structure Drive Behavioral and Ecological Variation in Endangered Coimbra Filho's Titi MonkeysDocument11 pagesFood Availability, Plant Diversity, and Vegetation Structure Drive Behavioral and Ecological Variation in Endangered Coimbra Filho's Titi MonkeysPaula FessiaNo ratings yet
- The Book of Seeds: A Life-Size Guide to Six Hundred Species from Around the WorldFrom EverandThe Book of Seeds: A Life-Size Guide to Six Hundred Species from Around the WorldRating: 4 out of 5 stars4/5 (1)
- Colloidal Aspects of Ice Cream-A ReviewDocument11 pagesColloidal Aspects of Ice Cream-A ReviewYuan NingNo ratings yet
- C MCQDocument450 pagesC MCQRuchi Sharma100% (1)
- PLC s7 AhmadiDocument59 pagesPLC s7 Ahmadisina20795No ratings yet
- Jeep 105Document10 pagesJeep 105Ishan LakhwaniNo ratings yet
- Microcontroller TroubleshootingDocument6 pagesMicrocontroller TroubleshootingMuhammad ZamirNo ratings yet
- Velocium Announcement From Hittite Oct 2007Document4 pagesVelocium Announcement From Hittite Oct 2007fleksNo ratings yet
- Screening Machine RHEWUM WA 606d523ae0775Document2 pagesScreening Machine RHEWUM WA 606d523ae0775calvarez alvarezNo ratings yet
- 475 - Assignment 3Document2 pages475 - Assignment 3sherasaabNo ratings yet
- Flexible Disc Couplings: TB Wood's Form-FlexDocument44 pagesFlexible Disc Couplings: TB Wood's Form-FlexAbhijeet KeerNo ratings yet
- Database and Transactions Management System For A Smart Gym: Layyah Fitness CenterDocument15 pagesDatabase and Transactions Management System For A Smart Gym: Layyah Fitness CenterAbu jor GifariNo ratings yet
- Electric Motor Start-Run Capacitor Selection GuideDocument18 pagesElectric Motor Start-Run Capacitor Selection GuideScott100% (7)
- Transpression and Transtension ZonesDocument14 pagesTranspression and Transtension Zonessolapado100% (1)
- 3BDS009030R4001 CIO PROFIBUS DP Engineering and ConfiguratioDocument89 pages3BDS009030R4001 CIO PROFIBUS DP Engineering and ConfiguratioGilney FreitasNo ratings yet
- Bitg 1113012003Document31 pagesBitg 1113012003kelipbuayaNo ratings yet
- Line BalancingDocument9 pagesLine BalancingJignesh SoniNo ratings yet
- Angle of Internal Friction and STP N Value RelationshipDocument25 pagesAngle of Internal Friction and STP N Value RelationshipHaseeb Hassan Gondal100% (1)
- Fatigue Resistance of The Binder Jet Printed 17-4 Precipitation Hardened Martensitic Stainless SteelDocument13 pagesFatigue Resistance of The Binder Jet Printed 17-4 Precipitation Hardened Martensitic Stainless SteelFathia AlkelaeNo ratings yet
- Draft StatisticsDocument2 pagesDraft StatisticsDanica PeñaNo ratings yet
- DEA351 A-Series Lighting Control Panel CatalogDocument8 pagesDEA351 A-Series Lighting Control Panel CatalogUlises Hernandez SolisNo ratings yet
- Edexcel Past Paper Maths CalDocument24 pagesEdexcel Past Paper Maths CalNazrul IslamNo ratings yet
- Networker 9.1.X Recommendations, Training and Downloads For Customers and PartnersDocument3 pagesNetworker 9.1.X Recommendations, Training and Downloads For Customers and PartnersAnonymous v89cvANo ratings yet
- Reading Material 2 - College Logic and MathDocument6 pagesReading Material 2 - College Logic and Mathjohn paul PatronNo ratings yet
- Chapter 3Document60 pagesChapter 3Czyrra Lyn Dimapush FetalverNo ratings yet
- Synapse Neurobiology: Week 5 - Part 1Document27 pagesSynapse Neurobiology: Week 5 - Part 1bobobNo ratings yet
- Magnetic Properties of Materials DocsDocument6 pagesMagnetic Properties of Materials DocsJeterine AriasNo ratings yet
- Activacion Asa 8.4 en Gns3: - VNC None - Vga None - M 1024 - Icount Auto - Hdachs 980,16,32Document10 pagesActivacion Asa 8.4 en Gns3: - VNC None - Vga None - M 1024 - Icount Auto - Hdachs 980,16,32Anibal Velozo VelozoNo ratings yet
- Simulated AnnealingDocument11 pagesSimulated Annealinggabby209No ratings yet
- Collina 2009Document14 pagesCollina 2009Research1 amtdcNo ratings yet
- Assignment 12Document3 pagesAssignment 12MAGESH K SEC 2020No ratings yet





























































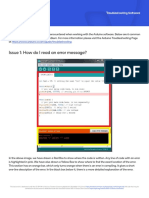
























Download as pdf or txt
You might also like
- Systems Architecture 7th Edition Burd Solutions Manual PDFDocument5 pagesSystems Architecture 7th Edition Burd Solutions Manual PDFa203025133No ratings yet
- Survival and Growth of Brazil-Nut Seedlings in Tree-Fall Gaps and Forest UnderstoryDocument10 pagesSurvival and Growth of Brazil-Nut Seedlings in Tree-Fall Gaps and Forest UnderstoryMarx Herrera MachacaNo ratings yet
- A Synopsis of The Rubiaceae of The States of Mato Grosso and Mato Grosso Do Sul, Brazil, With A Key To Genera, and A Preliminary Species ListDocument84 pagesA Synopsis of The Rubiaceae of The States of Mato Grosso and Mato Grosso Do Sul, Brazil, With A Key To Genera, and A Preliminary Species ListJúnior SenseyNo ratings yet
- Atlantic Forest To The All Americas: Biogeographical History and Divergence Times of Neotropical Ficus (Moraceae)Document51 pagesAtlantic Forest To The All Americas: Biogeographical History and Divergence Times of Neotropical Ficus (Moraceae)Francione GomesNo ratings yet
- Zuidema-Demografia de Nuez de Brazil Impactada Por El AprovechamientoDocument32 pagesZuidema-Demografia de Nuez de Brazil Impactada Por El AprovechamientoAlexis CentenoNo ratings yet
- Mares ErnstDocument20 pagesMares ErnstJoana FerreiraNo ratings yet
- Faunistic Analysis of Longhorn Beetles - Cerambycidae Coleoptera - in Cerrado and Atlantic Forest Areas Biodiversity Hotspots of BrazilDocument16 pagesFaunistic Analysis of Longhorn Beetles - Cerambycidae Coleoptera - in Cerrado and Atlantic Forest Areas Biodiversity Hotspots of BrazilMayre Ellen RodriguesNo ratings yet
- Papel de Los Mamíferos en Los Procesos de Dispersión y Depredación de Semillas de Mauritia Flexuosa (Arecaceae) en La..Document13 pagesPapel de Los Mamíferos en Los Procesos de Dispersión y Depredación de Semillas de Mauritia Flexuosa (Arecaceae) en La..Mariana Gutierrez MuneraNo ratings yet
- Lima Et Al. 2022 - NovaHedwigiaSemiaridchecklistDocument33 pagesLima Et Al. 2022 - NovaHedwigiaSemiaridchecklistJuliana LimaNo ratings yet
- Diet Specialisation in BirdsDocument17 pagesDiet Specialisation in BirdsИванАнато́лийФранкоNo ratings yet
- El Ecosistema de Dehesa Como ReservorioDocument13 pagesEl Ecosistema de Dehesa Como ReservorioIjoseangel BandinNo ratings yet
- Resturacion de IncendiosDocument9 pagesResturacion de IncendiosSalem Paco San MiguelNo ratings yet
- Biogeographic Patterns and Conservation in The South American Cerrado: A Tropical Savanna HotspotDocument12 pagesBiogeographic Patterns and Conservation in The South American Cerrado: A Tropical Savanna HotspotANDRES RICARDO LOPEZ PRECIADONo ratings yet
- A Comparison of The Wood Anatomy of 11 Species From Two Cerrado Habitats (Cerrado S.S. and Adjacent Gallery Forest) - Sonsin Et AlDocument20 pagesA Comparison of The Wood Anatomy of 11 Species From Two Cerrado Habitats (Cerrado S.S. and Adjacent Gallery Forest) - Sonsin Et Allihelih1No ratings yet
- ACFrOgDTOiHKN1mFw y7bSdQSLnMblq24oT1CulR6ZiJKiPERJ4g - CyHiovNdl AuNLd pXlWYtTXUny ZjmFgLdMJ51j2hUnRG3mCV0n3 - w4yZHMffhN7ngjTh - d08Ezh8Mv3SNY4GHwDwIbiKsDocument8 pagesACFrOgDTOiHKN1mFw y7bSdQSLnMblq24oT1CulR6ZiJKiPERJ4g - CyHiovNdl AuNLd pXlWYtTXUny ZjmFgLdMJ51j2hUnRG3mCV0n3 - w4yZHMffhN7ngjTh - d08Ezh8Mv3SNY4GHwDwIbiKsLaura MirandaNo ratings yet
- Providing Habitat For Native Mammals Through Understory Enhancement in Forestry PlantationsDocument5 pagesProviding Habitat For Native Mammals Through Understory Enhancement in Forestry Plantationsvalentina echeverriaNo ratings yet
- Janisovaal 2016 BullIAVSDocument17 pagesJanisovaal 2016 BullIAVSrangelbioNo ratings yet
- Goulart 2009Document9 pagesGoulart 2009Joana FerreiraNo ratings yet
- Non-Continuous Reproductive Phenology of Animal-Dispersed Species in Young Forest Restoration PlantingsDocument10 pagesNon-Continuous Reproductive Phenology of Animal-Dispersed Species in Young Forest Restoration PlantingsLeonardo UrruthNo ratings yet
- Bromelicolous Habit of Three Species of Rhinella Fitzinger, 1826 (Anura: Bufonidae) in Northeastern Brazil's Atlantic ForestDocument4 pagesBromelicolous Habit of Three Species of Rhinella Fitzinger, 1826 (Anura: Bufonidae) in Northeastern Brazil's Atlantic Forestvanessa e bruno becaciciNo ratings yet
- Leite Et Al 2013 - Foraging Behavior of Kaempfer's WoodpeckerDocument10 pagesLeite Et Al 2013 - Foraging Behavior of Kaempfer's WoodpeckerDianes Gomes MarcelinoNo ratings yet
- GiraldoValle CativoDocument20 pagesGiraldoValle CativoClaudia ParadaNo ratings yet
- Carlos A. Peres Luis C. Schiesari Claudia L. Dias-LemeDocument12 pagesCarlos A. Peres Luis C. Schiesari Claudia L. Dias-LemeItalo Treviño ZevallosNo ratings yet
- Galitti & Lovejoy - 2001 - The Future Atlantic Forest and Impact of LoggingDocument2 pagesGalitti & Lovejoy - 2001 - The Future Atlantic Forest and Impact of LoggingAllane SantosNo ratings yet
- Missouri Botanical Garden PressDocument34 pagesMissouri Botanical Garden PressAriadne Cristina De AntonioNo ratings yet
- The Brazilian Cerrado Vegetation and Threats To Its BiodiversityDocument8 pagesThe Brazilian Cerrado Vegetation and Threats To Its BiodiversityJackson RodriguesNo ratings yet
- Alvez-Valles Et Al. 2018 - Riqueza de Palmeiras Na AmazôniaDocument14 pagesAlvez-Valles Et Al. 2018 - Riqueza de Palmeiras Na AmazôniaGabriele FurtadoNo ratings yet
- Ecología Poblacional Del Ratón Peromyscus MexicanusDocument14 pagesEcología Poblacional Del Ratón Peromyscus MexicanusJuanNo ratings yet
- Plecevo Article 95601 en 1Document18 pagesPlecevo Article 95601 en 1Maurício GonçalvesNo ratings yet
- 1 PBDocument7 pages1 PBMylena NecoNo ratings yet
- 2 Macro PDFDocument22 pages2 Macro PDFLINA MARIA RODRIGUEZ RUIZNo ratings yet
- New Records of Macrofungi (Basidiomycota) For Colombia From A Tropical Dry ForestDocument22 pagesNew Records of Macrofungi (Basidiomycota) For Colombia From A Tropical Dry ForestLINA MARIA RODRIGUEZ RUIZNo ratings yet
- Caminha-Paiva Et Al 2022 - Fitossoc Canga QuartzitoDocument9 pagesCaminha-Paiva Et Al 2022 - Fitossoc Canga QuartzitoGeorgia SilvaNo ratings yet
- 2006-Viani, R.A.G. Et Al - Transference of Seedlings and Aloctone Young Individuals As Ecological Restoration MethodologyDocument26 pages2006-Viani, R.A.G. Et Al - Transference of Seedlings and Aloctone Young Individuals As Ecological Restoration MethodologyAdriano ItoNo ratings yet
- Linking Tropical Biology With Human DimensionsDocument5 pagesLinking Tropical Biology With Human Dimensionspabloribah7314No ratings yet
- 18157-Article Text-55730-1-10-20160202Document12 pages18157-Article Text-55730-1-10-20160202Lucìa SolerNo ratings yet
- What Do We Know About Cassis Tuberosa (Mollusca: Cassidae), A Heavily Exploited Marine Gastropod?Document14 pagesWhat Do We Know About Cassis Tuberosa (Mollusca: Cassidae), A Heavily Exploited Marine Gastropod?Murti HanafiNo ratings yet
- Artibeus Reservas Energéticas BHDocument8 pagesArtibeus Reservas Energéticas BHÉrica MunhozNo ratings yet
- Small Mammal Assemblages in A Disturbed Tropical LDocument10 pagesSmall Mammal Assemblages in A Disturbed Tropical LHeiner SaldañaNo ratings yet
- The Evolving Role of Bertholletia Excelsa in Amazonia: Contributing To Local Livelihoods and Forest ConservationDocument21 pagesThe Evolving Role of Bertholletia Excelsa in Amazonia: Contributing To Local Livelihoods and Forest ConservationMarx Herrera MachacaNo ratings yet
- Epifitas Da Pedra AzulDocument13 pagesEpifitas Da Pedra AzulAndre AssisNo ratings yet
- Edat Oliveres MilenariesDocument4 pagesEdat Oliveres MilenarieseltruferoNo ratings yet
- Fauna DarlynDocument14 pagesFauna DarlynDARLYN ASNIT LOPEZ JIMENEZNo ratings yet
- Machado 2021 Biogeography MoraceaeDocument13 pagesMachado 2021 Biogeography MoraceaeJéssica NayaraNo ratings yet
- Sinopsis Taxonómica, Ecológica y Etnobotánica Del Género Tachigali Aubl. (Leguminosae) en La Región Del Cusco, PerúDocument26 pagesSinopsis Taxonómica, Ecológica y Etnobotánica Del Género Tachigali Aubl. (Leguminosae) en La Región Del Cusco, PerúAnonymous my6XAlNo ratings yet
- Community Structure and Abundance of Small Rodents at The Wave Front of Agroforestry and Forest in Alto Beni, Bolivia - 2020Document10 pagesCommunity Structure and Abundance of Small Rodents at The Wave Front of Agroforestry and Forest in Alto Beni, Bolivia - 2020Abel Tome CaetanoNo ratings yet
- 1 s2.0 S0367253021001171 MainDocument12 pages1 s2.0 S0367253021001171 MainValeria CasiqueNo ratings yet
- Christianini 2018 Birdsfightforasinglecavity Rev Bras OrnitholDocument4 pagesChristianini 2018 Birdsfightforasinglecavity Rev Bras OrnitholjuanNo ratings yet
- Alcantara Etal 2018 Velloziaceae DiversificationDocument16 pagesAlcantara Etal 2018 Velloziaceae Diversificationdrinkto remenberNo ratings yet
- Forest Ecology and Management: A B A A C CDocument6 pagesForest Ecology and Management: A B A A C CJanaine RochaNo ratings yet
- Diversity and Seasonality of Scarabaeinae (Coleoptera Scarabaeidae) in Forest Fragments in Santa Maria, Rio Grande Do Sul, BrazilDocument19 pagesDiversity and Seasonality of Scarabaeinae (Coleoptera Scarabaeidae) in Forest Fragments in Santa Maria, Rio Grande Do Sul, BrazilEduardo ValentimNo ratings yet
- DownloadDocument9 pagesDownloadpaolospadera72No ratings yet
- Martins Et Al 2017 - Técnicas de Manejo de MelinisDocument9 pagesMartins Et Al 2017 - Técnicas de Manejo de MelinisHenrique SantosNo ratings yet
- Mammalia Firstrecordof Platyrrhinusfusciventris Chiroptera Phyllostomidaeforthe CaatingabiomeDocument6 pagesMammalia Firstrecordof Platyrrhinusfusciventris Chiroptera Phyllostomidaeforthe CaatingabiomeSara GNo ratings yet
- Diversidad Alfa y Beta de La Comunidad de Reptiles en El Complejo Cenagoso de Zapatosa, ColombiaDocument34 pagesDiversidad Alfa y Beta de La Comunidad de Reptiles en El Complejo Cenagoso de Zapatosa, ColombiaRaúl Carrillo CostaNo ratings yet
- Amburana CearensisDocument7 pagesAmburana CearensisAnthony GuerraNo ratings yet
- Tropical Rainforest Research PaperDocument5 pagesTropical Rainforest Research Papergz8qs4dn100% (1)
- 10.1515 - Mammalia 2023 0040Document7 pages10.1515 - Mammalia 2023 0040Hernani OliveiraNo ratings yet
- Ancient Transformation, Current Conservation: Traditional Forest Management On The Iriri River, Brazilian AmazoniaDocument15 pagesAncient Transformation, Current Conservation: Traditional Forest Management On The Iriri River, Brazilian AmazoniaMauricio TorresNo ratings yet
- Food Availability, Plant Diversity, and Vegetation Structure Drive Behavioral and Ecological Variation in Endangered Coimbra Filho's Titi MonkeysDocument11 pagesFood Availability, Plant Diversity, and Vegetation Structure Drive Behavioral and Ecological Variation in Endangered Coimbra Filho's Titi MonkeysPaula FessiaNo ratings yet
- The Book of Seeds: A Life-Size Guide to Six Hundred Species from Around the WorldFrom EverandThe Book of Seeds: A Life-Size Guide to Six Hundred Species from Around the WorldRating: 4 out of 5 stars4/5 (1)
- Colloidal Aspects of Ice Cream-A ReviewDocument11 pagesColloidal Aspects of Ice Cream-A ReviewYuan NingNo ratings yet
- C MCQDocument450 pagesC MCQRuchi Sharma100% (1)
- PLC s7 AhmadiDocument59 pagesPLC s7 Ahmadisina20795No ratings yet
- Jeep 105Document10 pagesJeep 105Ishan LakhwaniNo ratings yet
- Microcontroller TroubleshootingDocument6 pagesMicrocontroller TroubleshootingMuhammad ZamirNo ratings yet
- Velocium Announcement From Hittite Oct 2007Document4 pagesVelocium Announcement From Hittite Oct 2007fleksNo ratings yet
- Screening Machine RHEWUM WA 606d523ae0775Document2 pagesScreening Machine RHEWUM WA 606d523ae0775calvarez alvarezNo ratings yet
- 475 - Assignment 3Document2 pages475 - Assignment 3sherasaabNo ratings yet
- Flexible Disc Couplings: TB Wood's Form-FlexDocument44 pagesFlexible Disc Couplings: TB Wood's Form-FlexAbhijeet KeerNo ratings yet
- Database and Transactions Management System For A Smart Gym: Layyah Fitness CenterDocument15 pagesDatabase and Transactions Management System For A Smart Gym: Layyah Fitness CenterAbu jor GifariNo ratings yet
- Electric Motor Start-Run Capacitor Selection GuideDocument18 pagesElectric Motor Start-Run Capacitor Selection GuideScott100% (7)
- Transpression and Transtension ZonesDocument14 pagesTranspression and Transtension Zonessolapado100% (1)
- 3BDS009030R4001 CIO PROFIBUS DP Engineering and ConfiguratioDocument89 pages3BDS009030R4001 CIO PROFIBUS DP Engineering and ConfiguratioGilney FreitasNo ratings yet
- Bitg 1113012003Document31 pagesBitg 1113012003kelipbuayaNo ratings yet
- Line BalancingDocument9 pagesLine BalancingJignesh SoniNo ratings yet
- Angle of Internal Friction and STP N Value RelationshipDocument25 pagesAngle of Internal Friction and STP N Value RelationshipHaseeb Hassan Gondal100% (1)
- Fatigue Resistance of The Binder Jet Printed 17-4 Precipitation Hardened Martensitic Stainless SteelDocument13 pagesFatigue Resistance of The Binder Jet Printed 17-4 Precipitation Hardened Martensitic Stainless SteelFathia AlkelaeNo ratings yet
- Draft StatisticsDocument2 pagesDraft StatisticsDanica PeñaNo ratings yet
- DEA351 A-Series Lighting Control Panel CatalogDocument8 pagesDEA351 A-Series Lighting Control Panel CatalogUlises Hernandez SolisNo ratings yet
- Edexcel Past Paper Maths CalDocument24 pagesEdexcel Past Paper Maths CalNazrul IslamNo ratings yet
- Networker 9.1.X Recommendations, Training and Downloads For Customers and PartnersDocument3 pagesNetworker 9.1.X Recommendations, Training and Downloads For Customers and PartnersAnonymous v89cvANo ratings yet
- Reading Material 2 - College Logic and MathDocument6 pagesReading Material 2 - College Logic and Mathjohn paul PatronNo ratings yet
- Chapter 3Document60 pagesChapter 3Czyrra Lyn Dimapush FetalverNo ratings yet
- Synapse Neurobiology: Week 5 - Part 1Document27 pagesSynapse Neurobiology: Week 5 - Part 1bobobNo ratings yet
- Magnetic Properties of Materials DocsDocument6 pagesMagnetic Properties of Materials DocsJeterine AriasNo ratings yet
- Activacion Asa 8.4 en Gns3: - VNC None - Vga None - M 1024 - Icount Auto - Hdachs 980,16,32Document10 pagesActivacion Asa 8.4 en Gns3: - VNC None - Vga None - M 1024 - Icount Auto - Hdachs 980,16,32Anibal Velozo VelozoNo ratings yet
- Simulated AnnealingDocument11 pagesSimulated Annealinggabby209No ratings yet
- Collina 2009Document14 pagesCollina 2009Research1 amtdcNo ratings yet
- Assignment 12Document3 pagesAssignment 12MAGESH K SEC 2020No ratings yet