
Esteva Font2014 PDF
Esteva Font2014 PDF
You might also like
- Assignment 2 A Famous Contemporary LeaderDocument6 pagesAssignment 2 A Famous Contemporary Leaderfatin75% (4)
- Walk Through 3 Stars of DestinyDocument103 pagesWalk Through 3 Stars of DestinyLaura StanciuNo ratings yet
- WISC-IV 20adm ChecklistDocument20 pagesWISC-IV 20adm ChecklistJay Sun100% (2)
- Urea Transporters - 1Document7 pagesUrea Transporters - 1Steven Monge VindasNo ratings yet
- Blasi 2009Document8 pagesBlasi 2009veqmahwxqzzimuqymoNo ratings yet
- Permeabilidad Úrea FisioDocument8 pagesPermeabilidad Úrea FisioMaría Fe CasañoNo ratings yet
- Vasopressin Increases Phosphorylation of Ser84 and Ser486 in Slc14a2 Collecting Duct Urea TransportersDocument10 pagesVasopressin Increases Phosphorylation of Ser84 and Ser486 in Slc14a2 Collecting Duct Urea TransportersAlmas TNo ratings yet
- Intra Renal Cori CycleDocument21 pagesIntra Renal Cori Cyclesuman palNo ratings yet
- Polaritate HepatociteDocument45 pagesPolaritate HepatociteVetaNo ratings yet
- The Role of Transporters On Drug Therapy: NgatidjanDocument8 pagesThe Role of Transporters On Drug Therapy: NgatidjanMuhIrsalJumadilNo ratings yet
- Critical Reviews in Oral Biology & Medicine: Intracellular Transport and Secretion of Salivary ProteinsDocument20 pagesCritical Reviews in Oral Biology & Medicine: Intracellular Transport and Secretion of Salivary ProteinsNancy ArzateNo ratings yet
- Itagaki 2012Document6 pagesItagaki 2012Mohammad Nazmul IslamNo ratings yet
- Renal Drug Transporters and Their Signi Ficance in Drug - Drug InteractionsDocument11 pagesRenal Drug Transporters and Their Signi Ficance in Drug - Drug InteractionsHyosoo SungNo ratings yet
- 416 FullDocument9 pages416 FullFidia Hanan asystNo ratings yet
- Uapcr DiagnosticDocument8 pagesUapcr DiagnosticYA MAAPNo ratings yet
- Watford 2003Document9 pagesWatford 2003josetuiran992No ratings yet
- J - Pharmacol - Exp - Ther 2010 Watanabe 651 6 PDFDocument6 pagesJ - Pharmacol - Exp - Ther 2010 Watanabe 651 6 PDFMuriel NuñezNo ratings yet
- Hepatobiliary Excretion of Berberine: Pi-Lo Tsai and Tung-Hu TsaiDocument8 pagesHepatobiliary Excretion of Berberine: Pi-Lo Tsai and Tung-Hu TsaizebchemistNo ratings yet
- Chinkes1994 - Dog Pyruvate FlowDocument9 pagesChinkes1994 - Dog Pyruvate Flowreinafeng1No ratings yet
- The Role of Amino Acid Transporters in Inherited and Acquired DiseasesDocument19 pagesThe Role of Amino Acid Transporters in Inherited and Acquired DiseasesTuan TranNo ratings yet
- 1993 - The Vacuolar Membrane Protein Y-TIP Creates WaterDocument7 pages1993 - The Vacuolar Membrane Protein Y-TIP Creates WaterGiancarlo GHNo ratings yet
- Aquaporin Biology and Nervous System: Buffoli BarbaraDocument8 pagesAquaporin Biology and Nervous System: Buffoli BarbaraKhaled Abdel-saterNo ratings yet
- Uuo 1Document7 pagesUuo 1asaNo ratings yet
- Special Articles Serum Albumin: Marcus Murray Oratz Sidney S. SchreiberDocument17 pagesSpecial Articles Serum Albumin: Marcus Murray Oratz Sidney S. SchreiberTuấn AnhNo ratings yet
- DMD 106 010793 PDFDocument6 pagesDMD 106 010793 PDFTetteh JudeNo ratings yet
- Acetylated - Tubulin Is Reduced in Individuals With P - 2014 - Fertility and SteDocument13 pagesAcetylated - Tubulin Is Reduced in Individuals With P - 2014 - Fertility and Stelucifierop6No ratings yet
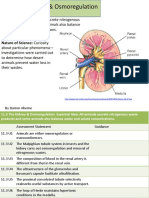
- 11.3 The Kidney & Osmoregulation: Essential Idea: All Animals Excrete NitrogenousDocument38 pages11.3 The Kidney & Osmoregulation: Essential Idea: All Animals Excrete NitrogenousmidahrazalNo ratings yet
- Articulo CientificoDocument17 pagesArticulo CientificoCarlos Rios PradaNo ratings yet
- Conte Camerino Neurochem Res 2004Document8 pagesConte Camerino Neurochem Res 2004pusch9728No ratings yet
- 1 Yosypiv2021 - Article - Renin-angiotensinSystemInMammaDocument11 pages1 Yosypiv2021 - Article - Renin-angiotensinSystemInMammaMuhammad Imran PanjwaniNo ratings yet
- Andre 1995 Membrane Transport Proteins SCDocument37 pagesAndre 1995 Membrane Transport Proteins SCAnaNo ratings yet
- Mechanisms of Disease: Review ArticleDocument11 pagesMechanisms of Disease: Review ArticleDámaris Barrueto MezaNo ratings yet
- Paper AdelaideDocument15 pagesPaper AdelaideSivle EvansNo ratings yet
- Are Mice, Rats, and Rabbits Good Models For Physiological, Pharmacological and Toxicological Studies in Humans?Document10 pagesAre Mice, Rats, and Rabbits Good Models For Physiological, Pharmacological and Toxicological Studies in Humans?Rifda LatifaNo ratings yet
- Evolutionary Analysis and Structural Insights in Brain Type Cytochrome P450 /aromatase of Stinging Catfish, Heteropneustes FossilisDocument27 pagesEvolutionary Analysis and Structural Insights in Brain Type Cytochrome P450 /aromatase of Stinging Catfish, Heteropneustes FossilisVerrell SeptianoNo ratings yet
- Urea and Ammonia Metabolism and The Control of Renal Nitrogen ExcretionDocument15 pagesUrea and Ammonia Metabolism and The Control of Renal Nitrogen ExcretionJames Cojab SacalNo ratings yet
- Role of Ion Channels in The Mechanism of ProteinuriaDocument10 pagesRole of Ion Channels in The Mechanism of ProteinuriaM Dwi SuprayogiNo ratings yet
- Maiuolo 2016Document7 pagesMaiuolo 2016okaNo ratings yet
- Serotonin L-Tryptophan and Tryptamine Are EffectivDocument7 pagesSerotonin L-Tryptophan and Tryptamine Are Effectivsyad deviskoNo ratings yet
- Cornichon 1Document16 pagesCornichon 1bryanmendietaNo ratings yet
- MolBiolEvol 2016 Wang Molbev msw191Document15 pagesMolBiolEvol 2016 Wang Molbev msw191zune153No ratings yet
- Joe 123Document19 pagesJoe 123mohammed fayedNo ratings yet
- 1 s2.0 S2214854X20300029 MainDocument11 pages1 s2.0 S2214854X20300029 MainFuad Fajar AkhlisNo ratings yet
- From in Silico To in Vitro: A Trip To Reveal Flavonoid Binding On The Rattus Norvegicus Kir6.1 ATP Sensitive Inward Rectifier Potassium ChannelDocument12 pagesFrom in Silico To in Vitro: A Trip To Reveal Flavonoid Binding On The Rattus Norvegicus Kir6.1 ATP Sensitive Inward Rectifier Potassium ChannelAurora PradoNo ratings yet
- Cold Spring Harb Perspect Biol-2017-Okamoto-cshperspect.a027839Document21 pagesCold Spring Harb Perspect Biol-2017-Okamoto-cshperspect.a027839crysraldixxNo ratings yet
- Sucrose Transporters of Higher PlantsDocument11 pagesSucrose Transporters of Higher PlantsRodica IonescuNo ratings yet
- Aquaporinas e Osmose PDFDocument11 pagesAquaporinas e Osmose PDFFábio Ximenes100% (1)
- 2004 The Epithelial Sodium ChannelDocument6 pages2004 The Epithelial Sodium ChannelyaritzaalviarezNo ratings yet
- EritropDocument9 pagesEritropSorin CalinNo ratings yet
- Pi Is 0085253815499661Document11 pagesPi Is 0085253815499661JundiNo ratings yet
- Histone Acetylation ReviewDocument7 pagesHistone Acetylation ReviewAshcharya RishiNo ratings yet
- 14_Schweizer et al 2014 - Structure and Function of Thyroid Hormone Plasma Membrane TransportersDocument11 pages14_Schweizer et al 2014 - Structure and Function of Thyroid Hormone Plasma Membrane TransporterswinterpruningNo ratings yet
- 2016.Zheng-Hai - Activating Nuclear Xenobiotic Receptors and Triggering ER Stress and Hepatic Cytochromes P450 Systems in Quails (Coturnix C-Coturnix) During Atrazine ExposureDocument10 pages2016.Zheng-Hai - Activating Nuclear Xenobiotic Receptors and Triggering ER Stress and Hepatic Cytochromes P450 Systems in Quails (Coturnix C-Coturnix) During Atrazine ExposureIasmin ResendeNo ratings yet
- Banta (1988) JCB 107, 1369-Vacuole Morphology of Vps MutantsDocument15 pagesBanta (1988) JCB 107, 1369-Vacuole Morphology of Vps MutantsDario FernándezNo ratings yet
- 4 DerlemeDocument6 pages4 DerlemeŞahin ŞahinNo ratings yet
- K Williams 1985 UP1 PNaSDocument5 pagesK Williams 1985 UP1 PNaSDolphingNo ratings yet
- Urinary Metabolic Phenotyping of WomenDocument9 pagesUrinary Metabolic Phenotyping of WomensusanaNo ratings yet
- tmpF9AC TMPDocument7 pagestmpF9AC TMPFrontiersNo ratings yet
- 10.1007@978 3 030 15950 411Document26 pages10.1007@978 3 030 15950 411Mabel Patricia Pereda SalgueroNo ratings yet
- Channels Uro BladderDocument12 pagesChannels Uro BladderKurniadi YusufNo ratings yet
- Bio HL Kidney SyllabusDocument2 pagesBio HL Kidney Syllabusniya.sokolova2006No ratings yet
- Renal Reflections: Exploring the Marvels of Nephrology: The Intricate Ballet of Kidney Functions UnveiledFrom EverandRenal Reflections: Exploring the Marvels of Nephrology: The Intricate Ballet of Kidney Functions UnveiledNo ratings yet
- Palash Agrawal ResumeDocument1 pagePalash Agrawal ResumesgbhuNo ratings yet
- Meditation For Beginners GuideDocument7 pagesMeditation For Beginners GuideEleven nameNo ratings yet
- INA827 Wide Supply Range, Rail-to-Rail Output Instrumentation Amplifier With A Minimum Gain of 5Document35 pagesINA827 Wide Supply Range, Rail-to-Rail Output Instrumentation Amplifier With A Minimum Gain of 5Sanjeev KumarNo ratings yet
- Departmentation or DepartmentalizationDocument1 pageDepartmentation or DepartmentalizationGulEFarisFarisNo ratings yet
- Aggregate Demand and Aggregate SupplyDocument34 pagesAggregate Demand and Aggregate SupplyGazal ReyazNo ratings yet
- The Effects of Pulsed Microwaves and Extra Low Frequency Electromagnetic Waves On Human BrainsDocument3 pagesThe Effects of Pulsed Microwaves and Extra Low Frequency Electromagnetic Waves On Human BrainsvalorecoragemNo ratings yet
- Atpl Theory Formulas PDFDocument64 pagesAtpl Theory Formulas PDFKumar AirplanefreakNo ratings yet
- Schedule of Evangelization - Noel AbanDocument2 pagesSchedule of Evangelization - Noel AbanJerome DesameroNo ratings yet
- Stairway To HeavenDocument17 pagesStairway To HeavenFrank gNo ratings yet
- Admin Law Project FinalDocument26 pagesAdmin Law Project FinalAnurag PrasharNo ratings yet
- Answers To Dragnet Abstract 1 & 2 PDFDocument9 pagesAnswers To Dragnet Abstract 1 & 2 PDFokotete evidenceNo ratings yet
- Survey, Investigation, Design and Estimation of Over Bridge at Budhheswar ChaurahaDocument100 pagesSurvey, Investigation, Design and Estimation of Over Bridge at Budhheswar ChaurahaManjeet SinghNo ratings yet
- Capacitive TransducerDocument21 pagesCapacitive Transducersourabhyadav14No ratings yet
- Stage 1 Name of The Students:: English IiiDocument6 pagesStage 1 Name of The Students:: English IiiRussell Najera RejonNo ratings yet
- Chapter 8 - Shikhandi: Princess Amba..... However That Was Another MatterDocument5 pagesChapter 8 - Shikhandi: Princess Amba..... However That Was Another MatteranjalibhatsNo ratings yet
- Microsoft Power Point - Organsiational Culture MSC Revised 1010Document6 pagesMicrosoft Power Point - Organsiational Culture MSC Revised 1010mattstewartis9589No ratings yet
- Website FeaturesDocument2 pagesWebsite Featuresapi-495718035No ratings yet
- Critical Essay PrintDocument4 pagesCritical Essay PrintAce Del RosarioNo ratings yet
- Middle School Lesson PlanDocument3 pagesMiddle School Lesson Planapi-339151467No ratings yet
- 1 - Page Law / RFBTDocument11 pages1 - Page Law / RFBTChloe SnowNo ratings yet
- 03 Answer Sheet Talking AboutDocument8 pages03 Answer Sheet Talking AboutQuinterito QuizNo ratings yet
- HHDocument226 pagesHHAstitva ChauhanNo ratings yet
- Events of Disputes AnswersDocument7 pagesEvents of Disputes AnswersMohamed AliNo ratings yet
- David PPT Exp Ch05Document76 pagesDavid PPT Exp Ch05Gracia PrimaNo ratings yet
- Introduction To Huawei UMTS Radio Network Dimensioning (RND)Document61 pagesIntroduction To Huawei UMTS Radio Network Dimensioning (RND)darkmesiahNo ratings yet
- PDFDocument3 pagesPDFMohammed AL-sharafiNo ratings yet
- (123doc - VN) - 41-De-Thi-Thu-Trac-Nhiem-Dai-Hoc-Tieng-Anh-Co-Dap-An PDFDocument248 pages(123doc - VN) - 41-De-Thi-Thu-Trac-Nhiem-Dai-Hoc-Tieng-Anh-Co-Dap-An PDFTrang NguyenNo ratings yet























































































Download as pdf or txt
You might also like
- Assignment 2 A Famous Contemporary LeaderDocument6 pagesAssignment 2 A Famous Contemporary Leaderfatin75% (4)
- Walk Through 3 Stars of DestinyDocument103 pagesWalk Through 3 Stars of DestinyLaura StanciuNo ratings yet
- WISC-IV 20adm ChecklistDocument20 pagesWISC-IV 20adm ChecklistJay Sun100% (2)
- Urea Transporters - 1Document7 pagesUrea Transporters - 1Steven Monge VindasNo ratings yet
- Blasi 2009Document8 pagesBlasi 2009veqmahwxqzzimuqymoNo ratings yet
- Permeabilidad Úrea FisioDocument8 pagesPermeabilidad Úrea FisioMaría Fe CasañoNo ratings yet
- Vasopressin Increases Phosphorylation of Ser84 and Ser486 in Slc14a2 Collecting Duct Urea TransportersDocument10 pagesVasopressin Increases Phosphorylation of Ser84 and Ser486 in Slc14a2 Collecting Duct Urea TransportersAlmas TNo ratings yet
- Intra Renal Cori CycleDocument21 pagesIntra Renal Cori Cyclesuman palNo ratings yet
- Polaritate HepatociteDocument45 pagesPolaritate HepatociteVetaNo ratings yet
- The Role of Transporters On Drug Therapy: NgatidjanDocument8 pagesThe Role of Transporters On Drug Therapy: NgatidjanMuhIrsalJumadilNo ratings yet
- Critical Reviews in Oral Biology & Medicine: Intracellular Transport and Secretion of Salivary ProteinsDocument20 pagesCritical Reviews in Oral Biology & Medicine: Intracellular Transport and Secretion of Salivary ProteinsNancy ArzateNo ratings yet
- Itagaki 2012Document6 pagesItagaki 2012Mohammad Nazmul IslamNo ratings yet
- Renal Drug Transporters and Their Signi Ficance in Drug - Drug InteractionsDocument11 pagesRenal Drug Transporters and Their Signi Ficance in Drug - Drug InteractionsHyosoo SungNo ratings yet
- 416 FullDocument9 pages416 FullFidia Hanan asystNo ratings yet
- Uapcr DiagnosticDocument8 pagesUapcr DiagnosticYA MAAPNo ratings yet
- Watford 2003Document9 pagesWatford 2003josetuiran992No ratings yet
- J - Pharmacol - Exp - Ther 2010 Watanabe 651 6 PDFDocument6 pagesJ - Pharmacol - Exp - Ther 2010 Watanabe 651 6 PDFMuriel NuñezNo ratings yet
- Hepatobiliary Excretion of Berberine: Pi-Lo Tsai and Tung-Hu TsaiDocument8 pagesHepatobiliary Excretion of Berberine: Pi-Lo Tsai and Tung-Hu TsaizebchemistNo ratings yet
- Chinkes1994 - Dog Pyruvate FlowDocument9 pagesChinkes1994 - Dog Pyruvate Flowreinafeng1No ratings yet
- The Role of Amino Acid Transporters in Inherited and Acquired DiseasesDocument19 pagesThe Role of Amino Acid Transporters in Inherited and Acquired DiseasesTuan TranNo ratings yet
- 1993 - The Vacuolar Membrane Protein Y-TIP Creates WaterDocument7 pages1993 - The Vacuolar Membrane Protein Y-TIP Creates WaterGiancarlo GHNo ratings yet
- Aquaporin Biology and Nervous System: Buffoli BarbaraDocument8 pagesAquaporin Biology and Nervous System: Buffoli BarbaraKhaled Abdel-saterNo ratings yet
- Uuo 1Document7 pagesUuo 1asaNo ratings yet
- Special Articles Serum Albumin: Marcus Murray Oratz Sidney S. SchreiberDocument17 pagesSpecial Articles Serum Albumin: Marcus Murray Oratz Sidney S. SchreiberTuấn AnhNo ratings yet
- DMD 106 010793 PDFDocument6 pagesDMD 106 010793 PDFTetteh JudeNo ratings yet
- Acetylated - Tubulin Is Reduced in Individuals With P - 2014 - Fertility and SteDocument13 pagesAcetylated - Tubulin Is Reduced in Individuals With P - 2014 - Fertility and Stelucifierop6No ratings yet
- 11.3 The Kidney & Osmoregulation: Essential Idea: All Animals Excrete NitrogenousDocument38 pages11.3 The Kidney & Osmoregulation: Essential Idea: All Animals Excrete NitrogenousmidahrazalNo ratings yet
- Articulo CientificoDocument17 pagesArticulo CientificoCarlos Rios PradaNo ratings yet
- Conte Camerino Neurochem Res 2004Document8 pagesConte Camerino Neurochem Res 2004pusch9728No ratings yet
- 1 Yosypiv2021 - Article - Renin-angiotensinSystemInMammaDocument11 pages1 Yosypiv2021 - Article - Renin-angiotensinSystemInMammaMuhammad Imran PanjwaniNo ratings yet
- Andre 1995 Membrane Transport Proteins SCDocument37 pagesAndre 1995 Membrane Transport Proteins SCAnaNo ratings yet
- Mechanisms of Disease: Review ArticleDocument11 pagesMechanisms of Disease: Review ArticleDámaris Barrueto MezaNo ratings yet
- Paper AdelaideDocument15 pagesPaper AdelaideSivle EvansNo ratings yet
- Are Mice, Rats, and Rabbits Good Models For Physiological, Pharmacological and Toxicological Studies in Humans?Document10 pagesAre Mice, Rats, and Rabbits Good Models For Physiological, Pharmacological and Toxicological Studies in Humans?Rifda LatifaNo ratings yet
- Evolutionary Analysis and Structural Insights in Brain Type Cytochrome P450 /aromatase of Stinging Catfish, Heteropneustes FossilisDocument27 pagesEvolutionary Analysis and Structural Insights in Brain Type Cytochrome P450 /aromatase of Stinging Catfish, Heteropneustes FossilisVerrell SeptianoNo ratings yet
- Urea and Ammonia Metabolism and The Control of Renal Nitrogen ExcretionDocument15 pagesUrea and Ammonia Metabolism and The Control of Renal Nitrogen ExcretionJames Cojab SacalNo ratings yet
- Role of Ion Channels in The Mechanism of ProteinuriaDocument10 pagesRole of Ion Channels in The Mechanism of ProteinuriaM Dwi SuprayogiNo ratings yet
- Maiuolo 2016Document7 pagesMaiuolo 2016okaNo ratings yet
- Serotonin L-Tryptophan and Tryptamine Are EffectivDocument7 pagesSerotonin L-Tryptophan and Tryptamine Are Effectivsyad deviskoNo ratings yet
- Cornichon 1Document16 pagesCornichon 1bryanmendietaNo ratings yet
- MolBiolEvol 2016 Wang Molbev msw191Document15 pagesMolBiolEvol 2016 Wang Molbev msw191zune153No ratings yet
- Joe 123Document19 pagesJoe 123mohammed fayedNo ratings yet
- 1 s2.0 S2214854X20300029 MainDocument11 pages1 s2.0 S2214854X20300029 MainFuad Fajar AkhlisNo ratings yet
- From in Silico To in Vitro: A Trip To Reveal Flavonoid Binding On The Rattus Norvegicus Kir6.1 ATP Sensitive Inward Rectifier Potassium ChannelDocument12 pagesFrom in Silico To in Vitro: A Trip To Reveal Flavonoid Binding On The Rattus Norvegicus Kir6.1 ATP Sensitive Inward Rectifier Potassium ChannelAurora PradoNo ratings yet
- Cold Spring Harb Perspect Biol-2017-Okamoto-cshperspect.a027839Document21 pagesCold Spring Harb Perspect Biol-2017-Okamoto-cshperspect.a027839crysraldixxNo ratings yet
- Sucrose Transporters of Higher PlantsDocument11 pagesSucrose Transporters of Higher PlantsRodica IonescuNo ratings yet
- Aquaporinas e Osmose PDFDocument11 pagesAquaporinas e Osmose PDFFábio Ximenes100% (1)
- 2004 The Epithelial Sodium ChannelDocument6 pages2004 The Epithelial Sodium ChannelyaritzaalviarezNo ratings yet
- EritropDocument9 pagesEritropSorin CalinNo ratings yet
- Pi Is 0085253815499661Document11 pagesPi Is 0085253815499661JundiNo ratings yet
- Histone Acetylation ReviewDocument7 pagesHistone Acetylation ReviewAshcharya RishiNo ratings yet
- 14_Schweizer et al 2014 - Structure and Function of Thyroid Hormone Plasma Membrane TransportersDocument11 pages14_Schweizer et al 2014 - Structure and Function of Thyroid Hormone Plasma Membrane TransporterswinterpruningNo ratings yet
- 2016.Zheng-Hai - Activating Nuclear Xenobiotic Receptors and Triggering ER Stress and Hepatic Cytochromes P450 Systems in Quails (Coturnix C-Coturnix) During Atrazine ExposureDocument10 pages2016.Zheng-Hai - Activating Nuclear Xenobiotic Receptors and Triggering ER Stress and Hepatic Cytochromes P450 Systems in Quails (Coturnix C-Coturnix) During Atrazine ExposureIasmin ResendeNo ratings yet
- Banta (1988) JCB 107, 1369-Vacuole Morphology of Vps MutantsDocument15 pagesBanta (1988) JCB 107, 1369-Vacuole Morphology of Vps MutantsDario FernándezNo ratings yet
- 4 DerlemeDocument6 pages4 DerlemeŞahin ŞahinNo ratings yet
- K Williams 1985 UP1 PNaSDocument5 pagesK Williams 1985 UP1 PNaSDolphingNo ratings yet
- Urinary Metabolic Phenotyping of WomenDocument9 pagesUrinary Metabolic Phenotyping of WomensusanaNo ratings yet
- tmpF9AC TMPDocument7 pagestmpF9AC TMPFrontiersNo ratings yet
- 10.1007@978 3 030 15950 411Document26 pages10.1007@978 3 030 15950 411Mabel Patricia Pereda SalgueroNo ratings yet
- Channels Uro BladderDocument12 pagesChannels Uro BladderKurniadi YusufNo ratings yet
- Bio HL Kidney SyllabusDocument2 pagesBio HL Kidney Syllabusniya.sokolova2006No ratings yet
- Renal Reflections: Exploring the Marvels of Nephrology: The Intricate Ballet of Kidney Functions UnveiledFrom EverandRenal Reflections: Exploring the Marvels of Nephrology: The Intricate Ballet of Kidney Functions UnveiledNo ratings yet
- Palash Agrawal ResumeDocument1 pagePalash Agrawal ResumesgbhuNo ratings yet
- Meditation For Beginners GuideDocument7 pagesMeditation For Beginners GuideEleven nameNo ratings yet
- INA827 Wide Supply Range, Rail-to-Rail Output Instrumentation Amplifier With A Minimum Gain of 5Document35 pagesINA827 Wide Supply Range, Rail-to-Rail Output Instrumentation Amplifier With A Minimum Gain of 5Sanjeev KumarNo ratings yet
- Departmentation or DepartmentalizationDocument1 pageDepartmentation or DepartmentalizationGulEFarisFarisNo ratings yet
- Aggregate Demand and Aggregate SupplyDocument34 pagesAggregate Demand and Aggregate SupplyGazal ReyazNo ratings yet
- The Effects of Pulsed Microwaves and Extra Low Frequency Electromagnetic Waves On Human BrainsDocument3 pagesThe Effects of Pulsed Microwaves and Extra Low Frequency Electromagnetic Waves On Human BrainsvalorecoragemNo ratings yet
- Atpl Theory Formulas PDFDocument64 pagesAtpl Theory Formulas PDFKumar AirplanefreakNo ratings yet
- Schedule of Evangelization - Noel AbanDocument2 pagesSchedule of Evangelization - Noel AbanJerome DesameroNo ratings yet
- Stairway To HeavenDocument17 pagesStairway To HeavenFrank gNo ratings yet
- Admin Law Project FinalDocument26 pagesAdmin Law Project FinalAnurag PrasharNo ratings yet
- Answers To Dragnet Abstract 1 & 2 PDFDocument9 pagesAnswers To Dragnet Abstract 1 & 2 PDFokotete evidenceNo ratings yet
- Survey, Investigation, Design and Estimation of Over Bridge at Budhheswar ChaurahaDocument100 pagesSurvey, Investigation, Design and Estimation of Over Bridge at Budhheswar ChaurahaManjeet SinghNo ratings yet
- Capacitive TransducerDocument21 pagesCapacitive Transducersourabhyadav14No ratings yet
- Stage 1 Name of The Students:: English IiiDocument6 pagesStage 1 Name of The Students:: English IiiRussell Najera RejonNo ratings yet
- Chapter 8 - Shikhandi: Princess Amba..... However That Was Another MatterDocument5 pagesChapter 8 - Shikhandi: Princess Amba..... However That Was Another MatteranjalibhatsNo ratings yet
- Microsoft Power Point - Organsiational Culture MSC Revised 1010Document6 pagesMicrosoft Power Point - Organsiational Culture MSC Revised 1010mattstewartis9589No ratings yet
- Website FeaturesDocument2 pagesWebsite Featuresapi-495718035No ratings yet
- Critical Essay PrintDocument4 pagesCritical Essay PrintAce Del RosarioNo ratings yet
- Middle School Lesson PlanDocument3 pagesMiddle School Lesson Planapi-339151467No ratings yet
- 1 - Page Law / RFBTDocument11 pages1 - Page Law / RFBTChloe SnowNo ratings yet
- 03 Answer Sheet Talking AboutDocument8 pages03 Answer Sheet Talking AboutQuinterito QuizNo ratings yet
- HHDocument226 pagesHHAstitva ChauhanNo ratings yet
- Events of Disputes AnswersDocument7 pagesEvents of Disputes AnswersMohamed AliNo ratings yet
- David PPT Exp Ch05Document76 pagesDavid PPT Exp Ch05Gracia PrimaNo ratings yet
- Introduction To Huawei UMTS Radio Network Dimensioning (RND)Document61 pagesIntroduction To Huawei UMTS Radio Network Dimensioning (RND)darkmesiahNo ratings yet
- PDFDocument3 pagesPDFMohammed AL-sharafiNo ratings yet
- (123doc - VN) - 41-De-Thi-Thu-Trac-Nhiem-Dai-Hoc-Tieng-Anh-Co-Dap-An PDFDocument248 pages(123doc - VN) - 41-De-Thi-Thu-Trac-Nhiem-Dai-Hoc-Tieng-Anh-Co-Dap-An PDFTrang NguyenNo ratings yet