
Basics of Protein Degradation
Basics of Protein Degradation
You might also like
- Arun PPT - PPTX ..HUMAN REPRODUCTION FOR HSC Board Maharashtra State..Document65 pagesArun PPT - PPTX ..HUMAN REPRODUCTION FOR HSC Board Maharashtra State..arunmishra1986100% (1)
- Cell NursingDocument83 pagesCell NursingRocKyRiazNo ratings yet
- Cell Bio Library WorkDocument5 pagesCell Bio Library WorkJP SilvaNo ratings yet
- Lysosomes TheCell NCBIBookshelf - 1705678132615Document3 pagesLysosomes TheCell NCBIBookshelf - 1705678132615kalitutawa465No ratings yet
- Lysosomes: Semester - IiDocument14 pagesLysosomes: Semester - IiJyoti DasNo ratings yet
- BIOC 315 - Lecture - 6Document46 pagesBIOC 315 - Lecture - 6Kevin KipropNo ratings yet
- Human PhysiologyDocument35 pagesHuman PhysiologyGanesha Dattatraya Gaonkar CFALNo ratings yet
- EXAM 1 ObjectivesDocument9 pagesEXAM 1 ObjectivesNoslaidysNo ratings yet
- LysosomesDocument29 pagesLysosomesBhavana ReddyNo ratings yet
- 0-Introduction NotesDocument7 pages0-Introduction Notesjimmy.bousaba27No ratings yet
- Misfolded ProteinsDocument8 pagesMisfolded ProteinsAfaq AhmadNo ratings yet
- CH RPTDocument12 pagesCH RPTDTM FHKNo ratings yet
- Plant CellDocument8 pagesPlant CellIrish Nicole PacionNo ratings yet
- LysosomesDocument15 pagesLysosomesIqra MujeebNo ratings yet
- LisosomDocument3 pagesLisosomAnonymous O1xkZINNo ratings yet
- The CellDocument72 pagesThe CellsirimaanuelNo ratings yet
- EC10 Secretion Lysosomes Peroxisomes TextDocument8 pagesEC10 Secretion Lysosomes Peroxisomes TextSilvito94No ratings yet
- CropDocument11 pagesCropHall OwnNo ratings yet
- LO Week 7Document22 pagesLO Week 7Matthew ChristopherNo ratings yet
- Proteasome AssignmentDocument12 pagesProteasome AssignmentMaryam AhmadiNo ratings yet
- Module 3 - Lecture 3 - LectureDocument8 pagesModule 3 - Lecture 3 - LectureNi CaoNo ratings yet
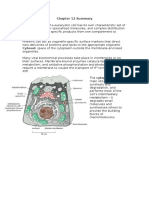
- Chapter 12 SummaryDocument7 pagesChapter 12 SummaryCharlotteNo ratings yet
- LysosomesDocument5 pagesLysosomespanigrahipuspanjali72No ratings yet
- Biology QuestionsDocument2 pagesBiology Questionsana sofiaNo ratings yet
- LYSOZOMEDocument6 pagesLYSOZOMETejas krishnakanthNo ratings yet
- Modification and Degradation of ProteinDocument6 pagesModification and Degradation of ProteinMuhammad Usman TahirNo ratings yet
- Partoza - JB, Concept Article - Endomembranesystem - DocxDocument4 pagesPartoza - JB, Concept Article - Endomembranesystem - DocxJenny PartozaNo ratings yet
- Cells, Organelles, Microscopy TechniquesDocument34 pagesCells, Organelles, Microscopy TechniquesJeffrey100% (1)
- Structural Biochemistry/Cell Organelles: Pictorial Representation of Organelles in A Typical Animal CellDocument8 pagesStructural Biochemistry/Cell Organelles: Pictorial Representation of Organelles in A Typical Animal CellPrincess Angie GonzalesNo ratings yet
- Phospholipids (Fats With Phosphorous Attached), Which at Body Temperature AreDocument7 pagesPhospholipids (Fats With Phosphorous Attached), Which at Body Temperature AreCmae VidadNo ratings yet
- Critical Review: Protein TurnoverDocument7 pagesCritical Review: Protein TurnoverSathish KumarNo ratings yet
- Nucleic AcidDocument3 pagesNucleic AciddspssmpNo ratings yet
- Biomed 04Document10 pagesBiomed 04api-3706483No ratings yet
- Cells and Their FunctionDocument4 pagesCells and Their FunctionMatika John PierreNo ratings yet
- LYSOSOMEDocument52 pagesLYSOSOMEDaniel Mendoza100% (1)
- The CellDocument41 pagesThe CellDANIELLA LOMA CAPONPONNo ratings yet
- Animal Physiology Theory Lec1Document24 pagesAnimal Physiology Theory Lec1ao868598No ratings yet
- Aging and Degradation of Proteins.-2Document3 pagesAging and Degradation of Proteins.-2carlottabovi28No ratings yet
- Endoplasmic ReticulumDocument12 pagesEndoplasmic ReticulumAzka AltafNo ratings yet
- Structure of The Cell Membrane (Plasma Membrane)Document6 pagesStructure of The Cell Membrane (Plasma Membrane)Umar AbdulmajeedNo ratings yet
- Lysosomes 160910093840Document32 pagesLysosomes 160910093840sekharurlaNo ratings yet
- Smooth and Granular Endoplasmic Reticulum RibosomesDocument26 pagesSmooth and Granular Endoplasmic Reticulum RibosomesArfiNo ratings yet
- Lysosomal LabizersDocument9 pagesLysosomal LabizersMalik OwaisNo ratings yet
- Physiology02 Cell PhysiologyDocument63 pagesPhysiology02 Cell PhysiologysmpoojasubashNo ratings yet
- The Cytoplasm: CytosolDocument5 pagesThe Cytoplasm: CytosolHariniNo ratings yet
- Cytosol 1Document20 pagesCytosol 1Swastu Nurul AzizahNo ratings yet
- Protein TargetingDocument10 pagesProtein TargetingdwigusmalawatiNo ratings yet
- Class 11 Biology Chapter 8 - Revision NotesDocument10 pagesClass 11 Biology Chapter 8 - Revision NotesHARKIRIT KAUR100% (1)
- Chapter Protein Folding and MisfoldingDocument17 pagesChapter Protein Folding and MisfoldingMazhar FarNo ratings yet
- Module 1 CellsDocument17 pagesModule 1 CellsLara C.No ratings yet
- 3.3 Tay-Sachs Niemann Pick Werkcollege 3Document6 pages3.3 Tay-Sachs Niemann Pick Werkcollege 3Josse BouwhuisNo ratings yet
- Mitochondria Week 7Document3 pagesMitochondria Week 7AamnaNo ratings yet
- Difference Between Lysosome and Peroxisome: March 2017Document10 pagesDifference Between Lysosome and Peroxisome: March 2017HNZY IAFSNo ratings yet
- Lysosomal Biology in CancerDocument16 pagesLysosomal Biology in CancerZi JianNo ratings yet
- Intro - The CellDocument31 pagesIntro - The CellJulius QuilapioNo ratings yet
- The Cell 1Document7 pagesThe Cell 1Eniola DaramolaNo ratings yet
- Biologi SelDocument3 pagesBiologi SelAmila MahriNo ratings yet
- NO.11 1. Golgi ApparatusDocument25 pagesNO.11 1. Golgi ApparatusEkaterina SurguladzeNo ratings yet
- AP Bio 6.4-6.6 OutlineDocument5 pagesAP Bio 6.4-6.6 OutlineShivali PatelNo ratings yet
- Lysosomal Function and Dysfunction: Mechanism and Disease: Orum Eview RticleDocument9 pagesLysosomal Function and Dysfunction: Mechanism and Disease: Orum Eview Rticleputri awaliah30No ratings yet
- HSC Biology Module 5Document21 pagesHSC Biology Module 5adneeniqbal259No ratings yet
- 2.3 Cellular Reproduction - NewDocument28 pages2.3 Cellular Reproduction - New23 JP NURFAEZZAH FAZRAHNI AKMADISNo ratings yet
- Biology 9700 Paper 2 - The Mitotic Cell CycleDocument43 pagesBiology 9700 Paper 2 - The Mitotic Cell CycleZainab AlziftawiNo ratings yet
- Development of Male Gametophyte (With Diagram) - Flowering Plants - Botany AwaisDocument4 pagesDevelopment of Male Gametophyte (With Diagram) - Flowering Plants - Botany AwaisAwais Kamran KhanNo ratings yet
- 2012exam2 With NotesDocument15 pages2012exam2 With NotesLillyNo ratings yet
- Science 5Document8 pagesScience 5100608No ratings yet
- Onion Root TipDocument5 pagesOnion Root TipGlaiza Nicole CuarteronNo ratings yet
- BMS532 L6 2020Document29 pagesBMS532 L6 2020zucc succNo ratings yet
- Cornell Notes - The Cell CycleDocument1 pageCornell Notes - The Cell Cycleapi-238397369No ratings yet
- MeiosisDocument23 pagesMeiosisJyothi GgNo ratings yet
- Class 12 Biology Pock BookDocument53 pagesClass 12 Biology Pock Bookbs.3136 VVS.No ratings yet
- Asexual Vs Sexual ReproductionDocument9 pagesAsexual Vs Sexual ReproductionEdenNo ratings yet
- DSE Biology Chapter 3C -Reproduction, growth and development 生殖、生長和發育Document37 pagesDSE Biology Chapter 3C -Reproduction, growth and development 生殖、生長和發育liNo ratings yet
- Transgenic AnimalsDocument9 pagesTransgenic Animals88AKKNo ratings yet
- Classification of PlantsDocument5 pagesClassification of PlantsodylorNo ratings yet
- One Gene One Enzyme TheoryDocument16 pagesOne Gene One Enzyme TheoryMuhammad ZakriaNo ratings yet
- Gene ExpressionDocument5 pagesGene ExpressionbenjaminlugahillaryNo ratings yet
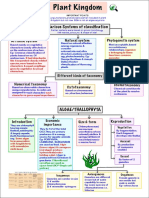
- 62e168e630d24 Plant Kingdom Mind MapDocument3 pages62e168e630d24 Plant Kingdom Mind MapDME MPonline100% (1)
- Student Exploration: MeiosisDocument8 pagesStudent Exploration: MeiosisGarnes TaahiraNo ratings yet
- Lec9 Cell Division and ChromosomeDocument29 pagesLec9 Cell Division and ChromosomeSITI BAZILAH BINTI BILAK KPM-GuruNo ratings yet
- Note 26 Jan 2024Document17 pagesNote 26 Jan 2024hs87s6smtnNo ratings yet
- Reproduction in OrganismsDocument7 pagesReproduction in Organisms.No ratings yet
- Reproduction in Animals Class 8Document7 pagesReproduction in Animals Class 8jahnavi poddar100% (1)
- Algae Structure and ReproductionDocument18 pagesAlgae Structure and ReproductionPrasanth Kumar S100% (1)
- 463-Article Text-2467-1-10-20200317Document14 pages463-Article Text-2467-1-10-20200317AOBO2OO32 Fadhila RamadhaniNo ratings yet
- DNA Mutation and RepairDocument8 pagesDNA Mutation and RepairShaimaa EmadNo ratings yet
- Modes of SpeciationDocument3 pagesModes of SpeciationRichard Balicat Jr.No ratings yet
- Bio Two Ses Chapter 6Document12 pagesBio Two Ses Chapter 6azizrafeeqNo ratings yet
- Grade 12 Topic Tests All CompressedDocument82 pagesGrade 12 Topic Tests All Compressedsamukeloandile4No ratings yet























































































Download as docx, pdf, or txt
You might also like
- Arun PPT - PPTX ..HUMAN REPRODUCTION FOR HSC Board Maharashtra State..Document65 pagesArun PPT - PPTX ..HUMAN REPRODUCTION FOR HSC Board Maharashtra State..arunmishra1986100% (1)
- Cell NursingDocument83 pagesCell NursingRocKyRiazNo ratings yet
- Cell Bio Library WorkDocument5 pagesCell Bio Library WorkJP SilvaNo ratings yet
- Lysosomes TheCell NCBIBookshelf - 1705678132615Document3 pagesLysosomes TheCell NCBIBookshelf - 1705678132615kalitutawa465No ratings yet
- Lysosomes: Semester - IiDocument14 pagesLysosomes: Semester - IiJyoti DasNo ratings yet
- BIOC 315 - Lecture - 6Document46 pagesBIOC 315 - Lecture - 6Kevin KipropNo ratings yet
- Human PhysiologyDocument35 pagesHuman PhysiologyGanesha Dattatraya Gaonkar CFALNo ratings yet
- EXAM 1 ObjectivesDocument9 pagesEXAM 1 ObjectivesNoslaidysNo ratings yet
- LysosomesDocument29 pagesLysosomesBhavana ReddyNo ratings yet
- 0-Introduction NotesDocument7 pages0-Introduction Notesjimmy.bousaba27No ratings yet
- Misfolded ProteinsDocument8 pagesMisfolded ProteinsAfaq AhmadNo ratings yet
- CH RPTDocument12 pagesCH RPTDTM FHKNo ratings yet
- Plant CellDocument8 pagesPlant CellIrish Nicole PacionNo ratings yet
- LysosomesDocument15 pagesLysosomesIqra MujeebNo ratings yet
- LisosomDocument3 pagesLisosomAnonymous O1xkZINNo ratings yet
- The CellDocument72 pagesThe CellsirimaanuelNo ratings yet
- EC10 Secretion Lysosomes Peroxisomes TextDocument8 pagesEC10 Secretion Lysosomes Peroxisomes TextSilvito94No ratings yet
- CropDocument11 pagesCropHall OwnNo ratings yet
- LO Week 7Document22 pagesLO Week 7Matthew ChristopherNo ratings yet
- Proteasome AssignmentDocument12 pagesProteasome AssignmentMaryam AhmadiNo ratings yet
- Module 3 - Lecture 3 - LectureDocument8 pagesModule 3 - Lecture 3 - LectureNi CaoNo ratings yet
- Chapter 12 SummaryDocument7 pagesChapter 12 SummaryCharlotteNo ratings yet
- LysosomesDocument5 pagesLysosomespanigrahipuspanjali72No ratings yet
- Biology QuestionsDocument2 pagesBiology Questionsana sofiaNo ratings yet
- LYSOZOMEDocument6 pagesLYSOZOMETejas krishnakanthNo ratings yet
- Modification and Degradation of ProteinDocument6 pagesModification and Degradation of ProteinMuhammad Usman TahirNo ratings yet
- Partoza - JB, Concept Article - Endomembranesystem - DocxDocument4 pagesPartoza - JB, Concept Article - Endomembranesystem - DocxJenny PartozaNo ratings yet
- Cells, Organelles, Microscopy TechniquesDocument34 pagesCells, Organelles, Microscopy TechniquesJeffrey100% (1)
- Structural Biochemistry/Cell Organelles: Pictorial Representation of Organelles in A Typical Animal CellDocument8 pagesStructural Biochemistry/Cell Organelles: Pictorial Representation of Organelles in A Typical Animal CellPrincess Angie GonzalesNo ratings yet
- Phospholipids (Fats With Phosphorous Attached), Which at Body Temperature AreDocument7 pagesPhospholipids (Fats With Phosphorous Attached), Which at Body Temperature AreCmae VidadNo ratings yet
- Critical Review: Protein TurnoverDocument7 pagesCritical Review: Protein TurnoverSathish KumarNo ratings yet
- Nucleic AcidDocument3 pagesNucleic AciddspssmpNo ratings yet
- Biomed 04Document10 pagesBiomed 04api-3706483No ratings yet
- Cells and Their FunctionDocument4 pagesCells and Their FunctionMatika John PierreNo ratings yet
- LYSOSOMEDocument52 pagesLYSOSOMEDaniel Mendoza100% (1)
- The CellDocument41 pagesThe CellDANIELLA LOMA CAPONPONNo ratings yet
- Animal Physiology Theory Lec1Document24 pagesAnimal Physiology Theory Lec1ao868598No ratings yet
- Aging and Degradation of Proteins.-2Document3 pagesAging and Degradation of Proteins.-2carlottabovi28No ratings yet
- Endoplasmic ReticulumDocument12 pagesEndoplasmic ReticulumAzka AltafNo ratings yet
- Structure of The Cell Membrane (Plasma Membrane)Document6 pagesStructure of The Cell Membrane (Plasma Membrane)Umar AbdulmajeedNo ratings yet
- Lysosomes 160910093840Document32 pagesLysosomes 160910093840sekharurlaNo ratings yet
- Smooth and Granular Endoplasmic Reticulum RibosomesDocument26 pagesSmooth and Granular Endoplasmic Reticulum RibosomesArfiNo ratings yet
- Lysosomal LabizersDocument9 pagesLysosomal LabizersMalik OwaisNo ratings yet
- Physiology02 Cell PhysiologyDocument63 pagesPhysiology02 Cell PhysiologysmpoojasubashNo ratings yet
- The Cytoplasm: CytosolDocument5 pagesThe Cytoplasm: CytosolHariniNo ratings yet
- Cytosol 1Document20 pagesCytosol 1Swastu Nurul AzizahNo ratings yet
- Protein TargetingDocument10 pagesProtein TargetingdwigusmalawatiNo ratings yet
- Class 11 Biology Chapter 8 - Revision NotesDocument10 pagesClass 11 Biology Chapter 8 - Revision NotesHARKIRIT KAUR100% (1)
- Chapter Protein Folding and MisfoldingDocument17 pagesChapter Protein Folding and MisfoldingMazhar FarNo ratings yet
- Module 1 CellsDocument17 pagesModule 1 CellsLara C.No ratings yet
- 3.3 Tay-Sachs Niemann Pick Werkcollege 3Document6 pages3.3 Tay-Sachs Niemann Pick Werkcollege 3Josse BouwhuisNo ratings yet
- Mitochondria Week 7Document3 pagesMitochondria Week 7AamnaNo ratings yet
- Difference Between Lysosome and Peroxisome: March 2017Document10 pagesDifference Between Lysosome and Peroxisome: March 2017HNZY IAFSNo ratings yet
- Lysosomal Biology in CancerDocument16 pagesLysosomal Biology in CancerZi JianNo ratings yet
- Intro - The CellDocument31 pagesIntro - The CellJulius QuilapioNo ratings yet
- The Cell 1Document7 pagesThe Cell 1Eniola DaramolaNo ratings yet
- Biologi SelDocument3 pagesBiologi SelAmila MahriNo ratings yet
- NO.11 1. Golgi ApparatusDocument25 pagesNO.11 1. Golgi ApparatusEkaterina SurguladzeNo ratings yet
- AP Bio 6.4-6.6 OutlineDocument5 pagesAP Bio 6.4-6.6 OutlineShivali PatelNo ratings yet
- Lysosomal Function and Dysfunction: Mechanism and Disease: Orum Eview RticleDocument9 pagesLysosomal Function and Dysfunction: Mechanism and Disease: Orum Eview Rticleputri awaliah30No ratings yet
- HSC Biology Module 5Document21 pagesHSC Biology Module 5adneeniqbal259No ratings yet
- 2.3 Cellular Reproduction - NewDocument28 pages2.3 Cellular Reproduction - New23 JP NURFAEZZAH FAZRAHNI AKMADISNo ratings yet
- Biology 9700 Paper 2 - The Mitotic Cell CycleDocument43 pagesBiology 9700 Paper 2 - The Mitotic Cell CycleZainab AlziftawiNo ratings yet
- Development of Male Gametophyte (With Diagram) - Flowering Plants - Botany AwaisDocument4 pagesDevelopment of Male Gametophyte (With Diagram) - Flowering Plants - Botany AwaisAwais Kamran KhanNo ratings yet
- 2012exam2 With NotesDocument15 pages2012exam2 With NotesLillyNo ratings yet
- Science 5Document8 pagesScience 5100608No ratings yet
- Onion Root TipDocument5 pagesOnion Root TipGlaiza Nicole CuarteronNo ratings yet
- BMS532 L6 2020Document29 pagesBMS532 L6 2020zucc succNo ratings yet
- Cornell Notes - The Cell CycleDocument1 pageCornell Notes - The Cell Cycleapi-238397369No ratings yet
- MeiosisDocument23 pagesMeiosisJyothi GgNo ratings yet
- Class 12 Biology Pock BookDocument53 pagesClass 12 Biology Pock Bookbs.3136 VVS.No ratings yet
- Asexual Vs Sexual ReproductionDocument9 pagesAsexual Vs Sexual ReproductionEdenNo ratings yet
- DSE Biology Chapter 3C -Reproduction, growth and development 生殖、生長和發育Document37 pagesDSE Biology Chapter 3C -Reproduction, growth and development 生殖、生長和發育liNo ratings yet
- Transgenic AnimalsDocument9 pagesTransgenic Animals88AKKNo ratings yet
- Classification of PlantsDocument5 pagesClassification of PlantsodylorNo ratings yet
- One Gene One Enzyme TheoryDocument16 pagesOne Gene One Enzyme TheoryMuhammad ZakriaNo ratings yet
- Gene ExpressionDocument5 pagesGene ExpressionbenjaminlugahillaryNo ratings yet
- 62e168e630d24 Plant Kingdom Mind MapDocument3 pages62e168e630d24 Plant Kingdom Mind MapDME MPonline100% (1)
- Student Exploration: MeiosisDocument8 pagesStudent Exploration: MeiosisGarnes TaahiraNo ratings yet
- Lec9 Cell Division and ChromosomeDocument29 pagesLec9 Cell Division and ChromosomeSITI BAZILAH BINTI BILAK KPM-GuruNo ratings yet
- Note 26 Jan 2024Document17 pagesNote 26 Jan 2024hs87s6smtnNo ratings yet
- Reproduction in OrganismsDocument7 pagesReproduction in Organisms.No ratings yet
- Reproduction in Animals Class 8Document7 pagesReproduction in Animals Class 8jahnavi poddar100% (1)
- Algae Structure and ReproductionDocument18 pagesAlgae Structure and ReproductionPrasanth Kumar S100% (1)
- 463-Article Text-2467-1-10-20200317Document14 pages463-Article Text-2467-1-10-20200317AOBO2OO32 Fadhila RamadhaniNo ratings yet
- DNA Mutation and RepairDocument8 pagesDNA Mutation and RepairShaimaa EmadNo ratings yet
- Modes of SpeciationDocument3 pagesModes of SpeciationRichard Balicat Jr.No ratings yet
- Bio Two Ses Chapter 6Document12 pagesBio Two Ses Chapter 6azizrafeeqNo ratings yet
- Grade 12 Topic Tests All CompressedDocument82 pagesGrade 12 Topic Tests All Compressedsamukeloandile4No ratings yet