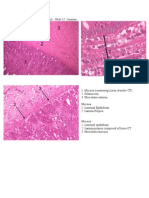
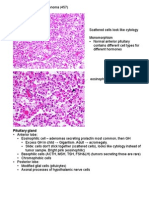
Download as pdf or txt
You might also like
- Medical Nutrition Therapy A Case Study Approach 5th Edition Nelms Solutions ManualDocument8 pagesMedical Nutrition Therapy A Case Study Approach 5th Edition Nelms Solutions Manualsophiechaurfqnz100% (34)
- 01a Using Anatomical LanguageDocument6 pages01a Using Anatomical Languageapi-3067625270% (1)
- CPC First TestDocument102 pagesCPC First TestChinnammal M100% (2)
- 4.thoracic WallDocument20 pages4.thoracic WallAmrith LordNo ratings yet
- Principal Parts of The Verb and Their FunctionsDocument2 pagesPrincipal Parts of The Verb and Their FunctionsJuanmiguel Ocampo Dion SchpNo ratings yet
- Cleavage, Blastula, Gastrula, NeurulaDocument41 pagesCleavage, Blastula, Gastrula, NeurulaHyenaNo ratings yet
- Perimeter Word ProblemsDocument3 pagesPerimeter Word ProblemsLayla AminNo ratings yet
- What Is FertilizationDocument8 pagesWhat Is Fertilizationapi-303065931No ratings yet
- 4) Describe The Structure and Functions of NeuronsDocument2 pages4) Describe The Structure and Functions of NeuronsKajolNo ratings yet
- Structure of NephronDocument8 pagesStructure of NephronPinkyFrancisNo ratings yet
- Grade 4 AnglesDocument4 pagesGrade 4 AnglesMohamed Mahmoud MobarakNo ratings yet
- BM 1-2-EcmDocument17 pagesBM 1-2-EcmSanthoshi Sadhanaa SankarNo ratings yet
- Other Disorders: Disorder Name: Congenital Adrenal Hyperplasia Acronym: CAHDocument14 pagesOther Disorders: Disorder Name: Congenital Adrenal Hyperplasia Acronym: CAHFriskaPieseshaNo ratings yet
- Muscle of Neck (Anatomy) PPT L.PPTX (Priti)Document21 pagesMuscle of Neck (Anatomy) PPT L.PPTX (Priti)Dj Manraj ProductionNo ratings yet
- Congenital Adrenal Hyperplasia (CAH)Document17 pagesCongenital Adrenal Hyperplasia (CAH)Rastu MaharthaNo ratings yet
- Chapter 4 Basic Geometrical IdeasDocument4 pagesChapter 4 Basic Geometrical Ideasraj mohanNo ratings yet
- The Process of FertilizationDocument16 pagesThe Process of FertilizationMarjorie ManlosaNo ratings yet
- Connective TissueDocument13 pagesConnective TissueHelena Allguter100% (3)
- 5 HypoxiaDocument42 pages5 HypoxiaShanzayNo ratings yet
- Plant EmbryogenesisDocument19 pagesPlant EmbryogenesisJosiahBriz-Rull100% (1)
- SpermatogenesisDocument24 pagesSpermatogenesisOmm KoliNo ratings yet
- Transport of Respiratory GasesDocument47 pagesTransport of Respiratory Gasessib786123No ratings yet
- 48 Hour ChickDocument13 pages48 Hour ChickJowi SalNo ratings yet
- BSC - 2019 - 1 - Anatomy and Physiology PDFDocument1 pageBSC - 2019 - 1 - Anatomy and Physiology PDFBasava VillanNo ratings yet
- 10 MM FrogDocument23 pages10 MM Frogfrances agulayNo ratings yet
- Embryo NotesDocument52 pagesEmbryo NotesSharona Avgush100% (1)
- Menstruatio N: Prepared By: Lirio, Riza D. BSN 2-1Document26 pagesMenstruatio N: Prepared By: Lirio, Riza D. BSN 2-1Eidyc_Etak_Sin_8542No ratings yet
- Ex234.1 Ascaris Eggz PDFDocument7 pagesEx234.1 Ascaris Eggz PDFCarlo MendozaNo ratings yet
- Respiratory System: StructureDocument29 pagesRespiratory System: StructureDr. Abir Ishtiaq100% (1)
- Lab 1. Population Genetics and Forces of EvolutionDocument8 pagesLab 1. Population Genetics and Forces of EvolutionKimberly YamNo ratings yet
- ANIMAL GROWTH AND DEVELOPMENT: The Tadpole (External Gill Stage)Document6 pagesANIMAL GROWTH AND DEVELOPMENT: The Tadpole (External Gill Stage)Junko TsukudaNo ratings yet
- Oo GenesisDocument20 pagesOo GenesisDinar Yudistira FirdausNo ratings yet
- The Species Concept-1Document5 pagesThe Species Concept-1O me khanNo ratings yet
- Urine Formation: Agnes R. Indrati Dept. of Clinical Pathology Univ. of PadjadjaranDocument36 pagesUrine Formation: Agnes R. Indrati Dept. of Clinical Pathology Univ. of PadjadjaranHafdzi MaulanaNo ratings yet
- 11 24 2006 MCB Chap - 16 - NDMCDocument16 pages11 24 2006 MCB Chap - 16 - NDMCapi-3696530No ratings yet
- MENSTRUATIONDocument13 pagesMENSTRUATIONHana-Lou TaquiquiNo ratings yet
- L7 - MeiosisDocument38 pagesL7 - MeiosisAl-Arqam OmarNo ratings yet
- AP Biology Outline - Chapter 5Document6 pagesAP Biology Outline - Chapter 5Omar LopezNo ratings yet
- Cycle CellDocument16 pagesCycle CellRohingya EnglishNo ratings yet
- Linear Algebra I: Course No. 100 221 Fall 2006 Michael StollDocument68 pagesLinear Algebra I: Course No. 100 221 Fall 2006 Michael StollDilip Kumar100% (1)
- 411 419Document9 pages411 419Afif MansorNo ratings yet
- HomeostasisDocument16 pagesHomeostasisDr M K GuptaNo ratings yet
- Extracellular Matrix (Ecm) : Dr. Naveen KumarDocument25 pagesExtracellular Matrix (Ecm) : Dr. Naveen Kumaraameena nadaNo ratings yet
- Anatomy and Blood Flow of LiverDocument45 pagesAnatomy and Blood Flow of Liverpukler1No ratings yet
- What Is Electrosurgery?Document4 pagesWhat Is Electrosurgery?RiyaNo ratings yet
- Embryology Embryology OF OF OF OF THE THE Respiratory Respiratory Respiratory Respiratory System SystemDocument40 pagesEmbryology Embryology OF OF OF OF THE THE Respiratory Respiratory Respiratory Respiratory System SystemChidera EmmanuelNo ratings yet
- Label The Brain Anatomy DiagramDocument2 pagesLabel The Brain Anatomy DiagramEmMa DHieyahNo ratings yet
- Complex Ion FormationDocument5 pagesComplex Ion FormationRendy Ahmad LubisNo ratings yet
- Problems On Normal DistributionDocument2 pagesProblems On Normal DistributionRavindra BabuNo ratings yet
- Genetics Study Guide AnswersDocument23 pagesGenetics Study Guide Answersapi-366982317No ratings yet
- Physiology CH 15 - Vascular Distensibility and FunctionDocument31 pagesPhysiology CH 15 - Vascular Distensibility and FunctionDaniel AdamsNo ratings yet
- 05 GastrulationDocument6 pages05 GastrulationcarlgangcaNo ratings yet
- Embryology of Digestive System1Document62 pagesEmbryology of Digestive System1Gina AnnisahNo ratings yet
- 2013 - Lower Limb-Fascial Compartments of Thigh - Student VersionDocument28 pages2013 - Lower Limb-Fascial Compartments of Thigh - Student Versionmarina_shawkyNo ratings yet
- 1 BloodDocument77 pages1 BloodVisaNathan100% (1)
- Introduction in Human Anatomy and PhysiologyDocument35 pagesIntroduction in Human Anatomy and Physiologyraul nino MoranNo ratings yet
- BIOLAB FROG Organ HistologyDocument7 pagesBIOLAB FROG Organ HistologyCloie Anne Rabinetas100% (1)
- Anatomy Notes LeydigDocument1 pageAnatomy Notes Leydigjed guicoNo ratings yet
- Early Development in BirdsDocument53 pagesEarly Development in BirdsMuqaddas NadeemNo ratings yet
- Dentogingival Junction (DGJ)Document8 pagesDentogingival Junction (DGJ)روان امير غني عبد عونNo ratings yet
- Urogenital SystemDocument5 pagesUrogenital Systemstudent10100No ratings yet
- Human FertilizationDocument3 pagesHuman Fertilizationtamalbio7No ratings yet
- 0000000000000000000000000000000Document1 page0000000000000000000000000000000KnjigeNo ratings yet
- This Document Was Uploaded by User and They Confirmed That They Have The Permission To Share ItDocument1 pageThis Document Was Uploaded by User and They Confirmed That They Have The Permission To Share ItKnjigeNo ratings yet
- ToxoplasmosisDocument27 pagesToxoplasmosisKnjigeNo ratings yet
- Procjena Zdravstvenih Potreba Model Životnoga Ciklusa: Vježba 3Document2 pagesProcjena Zdravstvenih Potreba Model Životnoga Ciklusa: Vježba 3KnjigeNo ratings yet
- ItalianToGo PDFDocument2 pagesItalianToGo PDFKnjigeNo ratings yet
- Ponašanje Profila Konstrukcije Dentalnog Mosta Prema Silama OpterećenjaDocument5 pagesPonašanje Profila Konstrukcije Dentalnog Mosta Prema Silama OpterećenjaKnjigeNo ratings yet
- Propranolol For HemangiomaDocument4 pagesPropranolol For HemangiomaJheelzy DatorNo ratings yet
- Mid-Term Examination PolygraphyDocument3 pagesMid-Term Examination PolygraphyMica EnriquezNo ratings yet
- Republic of The PhilippinesDocument3 pagesRepublic of The PhilippinesTroy JcNo ratings yet
- Curriculum of Plastic Surgery Egyptian BoardDocument34 pagesCurriculum of Plastic Surgery Egyptian BoardsenthiljayanthiNo ratings yet
- Meditation Can Change Your BrainDocument38 pagesMeditation Can Change Your BrainZena Chames Eddin100% (1)
- Tagar As MedicineDocument2 pagesTagar As MedicineNavotna ShuklaNo ratings yet
- Lydia HallDocument5 pagesLydia Hallbbasya eihyna100% (5)
- Biological-Psychology-reviewer-Copy-converted - PDF Version 1Document17 pagesBiological-Psychology-reviewer-Copy-converted - PDF Version 1Mark John DequitosNo ratings yet
- Introduction To Laboratory MedicineDocument308 pagesIntroduction To Laboratory MedicineJane JeriesNo ratings yet
- Intracranial Vascular Imaging: Pearls and Pitfalls: Jane J. Kim, MD, and Max Wintermark, MDDocument5 pagesIntracranial Vascular Imaging: Pearls and Pitfalls: Jane J. Kim, MD, and Max Wintermark, MDabowalid98No ratings yet
- Neurophysiology Learning ObjectivesDocument88 pagesNeurophysiology Learning ObjectivesBad BunnyNo ratings yet
- Storage - Emulated - 0 - Whatsapp - Media - WhatsApp Documents - NOTES CLASS 9 TISSUESDocument14 pagesStorage - Emulated - 0 - Whatsapp - Media - WhatsApp Documents - NOTES CLASS 9 TISSUESdubeymaya153No ratings yet
- Grade 10 Unit 3 - HomeostasisDocument24 pagesGrade 10 Unit 3 - HomeostasistameratNo ratings yet
- Name: Class: Fill in The Blank With Correct AnswerDocument1 pageName: Class: Fill in The Blank With Correct AnswerRok RuzNo ratings yet
- PhenanthrolineDocument3 pagesPhenanthrolineRega PermanaNo ratings yet
- Protein DigestionDocument198 pagesProtein DigestionLupuLoredanaNo ratings yet
- Coordination and Response in Humans: 12.6 Endocrine SystemDocument16 pagesCoordination and Response in Humans: 12.6 Endocrine Systemwienna1987No ratings yet
- Eng Hamid & Zakaria PDFDocument5 pagesEng Hamid & Zakaria PDFTiga TujuhNo ratings yet
- Final Notes PE2Document5 pagesFinal Notes PE2israel seraficaNo ratings yet
- Somatosensory System: G.Arvelandze N.MalashkhiaDocument26 pagesSomatosensory System: G.Arvelandze N.MalashkhialuckyNo ratings yet
- ProteinDocument16 pagesProteinGwen Cua50% (2)
- Endocrine Glands PathologyDocument8 pagesEndocrine Glands Pathologychrisp7No ratings yet
- Effect of Vitamin E On Thrombocytopenia in Dengue FeverDocument5 pagesEffect of Vitamin E On Thrombocytopenia in Dengue FeverBernadeth MeilyNo ratings yet
- ISOFLURANE ArroganteDocument2 pagesISOFLURANE ArroganteMheg ArroganteNo ratings yet
- Entropy Quick Guide - JB44337XX - Nov4 PDFDocument4 pagesEntropy Quick Guide - JB44337XX - Nov4 PDFElena CracNo ratings yet
- CVD NCPDocument3 pagesCVD NCPpromilokidNo ratings yet
- 1 Intro To HematologyDocument2 pages1 Intro To HematologyAldrin ChrisNo ratings yet
- Approach To Cyanosis in NewbornDocument28 pagesApproach To Cyanosis in NewbornbidisNo ratings yet