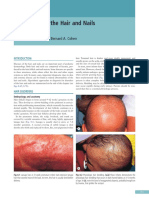
Download as pdf or txt
You might also like
- Beauty Salon TBDocument40 pagesBeauty Salon TBJoanna Smithson83% (12)
- Magical Hair - LeachDocument19 pagesMagical Hair - LeachSuranja Lakmal Perera100% (2)
- Androgenic Alopecia Finished ThesisDocument73 pagesAndrogenic Alopecia Finished ThesisAkshay R Aiyar100% (2)
- Lecture Notes On Dermatology PDFDocument201 pagesLecture Notes On Dermatology PDFmaskmaz100% (1)
- ALOPECIADocument90 pagesALOPECIAluckyswiss7776848No ratings yet
- Hair Follicle, Functions, Diseases, A Simple Guide To The Condition, Diagnosis, Treatment And Related ConditionsFrom EverandHair Follicle, Functions, Diseases, A Simple Guide To The Condition, Diagnosis, Treatment And Related ConditionsNo ratings yet
- Aboriginal History V35Document264 pagesAboriginal History V35candvbks100% (1)
- The Biology of Hair Follicles: New England Journal of Medicine September 1999Document8 pagesThe Biology of Hair Follicles: New England Journal of Medicine September 1999Henny KoleworaNo ratings yet
- Biology Hair 1999Document7 pagesBiology Hair 1999Ilse PalaciosNo ratings yet
- Male Pattern Androgenetic AlopeciaDocument6 pagesMale Pattern Androgenetic AlopeciaYasser Yousif Al-ahmadyNo ratings yet
- Hair Diseases: AnatomyDocument36 pagesHair Diseases: AnatomyMasithaNo ratings yet
- Presentation Electri in TrichologyDocument69 pagesPresentation Electri in TrichologycarmenNo ratings yet
- Dr. Dixit-MS 2nd ProofDocument22 pagesDr. Dixit-MS 2nd ProofGaneshKumarNo ratings yet
- Biology of The Hair Follicle: The Basics: Karoline Krause, MD, and Kerstin Foitzik, MDDocument9 pagesBiology of The Hair Follicle: The Basics: Karoline Krause, MD, and Kerstin Foitzik, MDSuzana PoloncaNo ratings yet
- Whiting 2001Document6 pagesWhiting 2001Nur ElidaNo ratings yet
- 12 Harish K-Rajesh R-Yasar-Arafath A-Ramzan-Beevi MDocument15 pages12 Harish K-Rajesh R-Yasar-Arafath A-Ramzan-Beevi MMichaelNo ratings yet
- The Diagnosis and Treatment of Hair PDFDocument11 pagesThe Diagnosis and Treatment of Hair PDFSpica AdharaNo ratings yet
- Disorders of HairDocument5 pagesDisorders of Hairعبدالعزيز احمد علي عتشNo ratings yet
- NEJM199909233411307Document10 pagesNEJM199909233411307evelynemayNo ratings yet
- Estudio XL HAIR - Alopecia AreataDocument32 pagesEstudio XL HAIR - Alopecia AreataIrina KucevichuteNo ratings yet
- The Biology of Hair Diversity: Review ArticleDocument8 pagesThe Biology of Hair Diversity: Review ArticleFernandaNo ratings yet
- Steroidify Hair Loss GuideDocument13 pagesSteroidify Hair Loss GuidefizzNo ratings yet
- Анатом. И Физиолог. ВолосаDocument17 pagesАнатом. И Физиолог. Волосаруслан барчевскийNo ratings yet
- THE Biology of Hair Care: Dermatologic Aspects of CosmeticsDocument8 pagesTHE Biology of Hair Care: Dermatologic Aspects of CosmeticsSiska B SiregarNo ratings yet
- Mary Clare Higgins-Chen, MD, MPH Donna Windish, MD, MPH: Alopecia Week 25Document9 pagesMary Clare Higgins-Chen, MD, MPH Donna Windish, MD, MPH: Alopecia Week 25omegasauron0gmailcomNo ratings yet
- Bender 2013Document29 pagesBender 2013Jesus Vargas GalindoNo ratings yet
- Article 0011 44 50Document7 pagesArticle 0011 44 50apj abdulNo ratings yet
- Physical Means of Treating Unwanted Hair: R W & T S. ADocument10 pagesPhysical Means of Treating Unwanted Hair: R W & T S. Adipika awindaNo ratings yet
- The Hair Follicle EnigmaDocument6 pagesThe Hair Follicle EnigmaLorena AcevedoNo ratings yet
- Articlehu 77Document14 pagesArticlehu 77Matheus De oliveiraNo ratings yet
- Childrens Hair LossDocument8 pagesChildrens Hair Lossdrmurad321No ratings yet
- Hair BiologyDocument16 pagesHair BiologyTakis VasilopoulosNo ratings yet
- Anatomy and Physiology of HairDocument16 pagesAnatomy and Physiology of HairLaura CaputoNo ratings yet
- Hair Fall TreatmentDocument11 pagesHair Fall TreatmentpankajkhakareNo ratings yet
- Fish N Chicks Models For Auditory Hair Cell RegenerationDocument4 pagesFish N Chicks Models For Auditory Hair Cell RegenerationTapan K. NayakNo ratings yet
- Harrison Sinclairhairstylingl-2003-Journal of Cosmetic DermatologyDocument7 pagesHarrison Sinclairhairstylingl-2003-Journal of Cosmetic Dermatologyلمياء لعلاج وفرد الشعرNo ratings yet
- Introduction To Hair Development: Duane P. HarlandDocument8 pagesIntroduction To Hair Development: Duane P. HarlandAlexandra DobrescuNo ratings yet
- The Wonder of Herbs To Treat-AlopeciaDocument7 pagesThe Wonder of Herbs To Treat-AlopeciaLuca JohnNo ratings yet
- Alopecia PDFDocument36 pagesAlopecia PDFFelp ScholzNo ratings yet
- 2002 - Millar - Molecular Mechanism Regulating Hair Follicle DevelopmentDocument10 pages2002 - Millar - Molecular Mechanism Regulating Hair Follicle DevelopmentalexantonjazzNo ratings yet
- Anagen EffluviumDocument9 pagesAnagen EffluviumMariana ÁvilaNo ratings yet
- Healthy Hair: What Is It?: Rodney D. SinclairDocument4 pagesHealthy Hair: What Is It?: Rodney D. SinclairDavid51No ratings yet
- Systematic Approach To Hair Loss in Women: Ralph M. TrüebDocument15 pagesSystematic Approach To Hair Loss in Women: Ralph M. TrüebErika Alvarez IbarraNo ratings yet
- Male Patternbaldness EllisDocument8 pagesMale Patternbaldness ElliszmahfudzNo ratings yet
- Assessment of Hair and Scalp Disorders: Antonella Tosti and John GrayDocument5 pagesAssessment of Hair and Scalp Disorders: Antonella Tosti and John GrayDavid51No ratings yet
- Ms 2007 Derm Norhan Ali HeiderDocument5 pagesMs 2007 Derm Norhan Ali HeiderAnnisa DarmawahyuniNo ratings yet
- Diseases of The Skin AppendagesDocument42 pagesDiseases of The Skin AppendagesEnverieNo ratings yet
- Hair Loss Common Causes and TreatmentDocument8 pagesHair Loss Common Causes and TreatmentDesireé González ArzolayNo ratings yet
- AlopeciaDocument90 pagesAlopeciasmbawasainiNo ratings yet
- Lesser Known Homoeopathic Medicines of Alopecia Areata.20200718115446Document9 pagesLesser Known Homoeopathic Medicines of Alopecia Areata.20200718115446BruntNo ratings yet
- Sinclair 2001Document12 pagesSinclair 2001lisnallNo ratings yet
- Livro de Remédios NaturaisDocument8 pagesLivro de Remédios NaturaisJeff SilvaNo ratings yet
- Androgenetic Alopecia: January 2015Document14 pagesAndrogenetic Alopecia: January 2015MFajri irfanNo ratings yet
- Hair Restoration Surgery: The State of The ArtDocument24 pagesHair Restoration Surgery: The State of The ArtMehmet Sürmeli100% (1)
- Hairbiology: Growth and PigmentationDocument10 pagesHairbiology: Growth and PigmentationAlexandra DobrescuNo ratings yet
- amazing studyDocument19 pagesamazing studynatasjlpNo ratings yet
- Management of Alopecia Areata in ChildrenDocument22 pagesManagement of Alopecia Areata in ChildrenAnita MarakNo ratings yet
- Hair Restoration Update: by DR - Tarek Ahmed Said Lecturer of Plastic Surgery, Cairo UniversityDocument27 pagesHair Restoration Update: by DR - Tarek Ahmed Said Lecturer of Plastic Surgery, Cairo UniversityHamid Reza TeymouriNo ratings yet
- Androgenic Alopecia For TehranDocument50 pagesAndrogenic Alopecia For TehranLuis Soriano MitraNo ratings yet
- 8 - Hair Disorders (Updated)Document14 pages8 - Hair Disorders (Updated)haytham aliNo ratings yet
- Annular Alopecia Areata: Report of Two CasesDocument4 pagesAnnular Alopecia Areata: Report of Two CasesZAHYRA MARÍA VALDERRAMA POLANIANo ratings yet
- Understanding Causes of Hair Loss in Women 2021Document4 pagesUnderstanding Causes of Hair Loss in Women 2021maat1No ratings yet
- Haircut TechniquesDocument2 pagesHaircut TechniquesAlyanna TanNo ratings yet
- Narcisse March 2006Document51 pagesNarcisse March 2006Carolyn Samuels0% (1)
- LESSON PLAN - Describing-PeopleDocument4 pagesLESSON PLAN - Describing-PeopleWnhannan MohamadNo ratings yet
- Genotype and Phenotype ActivityDocument3 pagesGenotype and Phenotype ActivityAri LimNo ratings yet
- Detumescence Therapy of Human Scalp For Natural Hair Regrowth 2155 9554.1000138Document5 pagesDetumescence Therapy of Human Scalp For Natural Hair Regrowth 2155 9554.1000138forex500No ratings yet
- Describing People VocabularyDocument3 pagesDescribing People VocabularyMarino GarciaNo ratings yet
- HaircuttingDocument15 pagesHaircuttingMary Rhea R Ileto100% (2)
- Tercer Unidad 3Document12 pagesTercer Unidad 3gbrielaNo ratings yet
- ST Mary S Clinic Booklet 1 0Document52 pagesST Mary S Clinic Booklet 1 0Lionel FarrugiaNo ratings yet
- Colourless Go Blonde InstructionsDocument2 pagesColourless Go Blonde InstructionsJanet SimovicNo ratings yet
- Diet For Healthy HairDocument4 pagesDiet For Healthy HairAsif IqbalNo ratings yet
- MHE 13. Bulaquiña (Unit 5)Document4 pagesMHE 13. Bulaquiña (Unit 5)SHEILA MAE BULAQUIÑANo ratings yet
- CGM ExplainedDocument7 pagesCGM ExplainedRomielle ArnaldoNo ratings yet
- Bingo Body PartsDocument22 pagesBingo Body PartsPao PerezNo ratings yet
- TELOGEN EFFLUVIUM (A Type of Hair Loss) : What Are The Aims of This Leaflet?Document4 pagesTELOGEN EFFLUVIUM (A Type of Hair Loss) : What Are The Aims of This Leaflet?Surendra KumarNo ratings yet
- Glándulas Prepuciales (Tyson)Document5 pagesGlándulas Prepuciales (Tyson)AlejandroVeraVargasNo ratings yet
- Newsletter End of Summer TermDocument8 pagesNewsletter End of Summer TermisleworthsyonNo ratings yet
- List I No Cosmetic oDocument32 pagesList I No Cosmetic oFarrensyah PutraNo ratings yet
- OthersDocument4 pagesOthersalankrutha devuNo ratings yet
- Autodesk Maya Digital Hair SeminarDocument61 pagesAutodesk Maya Digital Hair Seminartlo136No ratings yet
- Desert Daughter - HebrewDocument24 pagesDesert Daughter - HebrewIsrael ExporterNo ratings yet
- A 6-Month, Randomized, Double-Blind, Placebo-Controlled Study Evaluating The Ability of A Marine Complex Supplement To Promote Hair Growth in Men With Thinning HairDocument9 pagesA 6-Month, Randomized, Double-Blind, Placebo-Controlled Study Evaluating The Ability of A Marine Complex Supplement To Promote Hair Growth in Men With Thinning HairDimas PrajagoptaNo ratings yet
- 20 Recipes For Homemade Hair Growth TreatmentsDocument41 pages20 Recipes For Homemade Hair Growth Treatmentslara-usui100% (2)
- Beauty CultureDocument410 pagesBeauty CultureAndromeda MNo ratings yet
- Fischer Et Al-2008-Journal of Pineal ResearchDocument15 pagesFischer Et Al-2008-Journal of Pineal ResearchgeoaislaNo ratings yet