Download as pdf or txt
You might also like
- ACT Science Stu 2014 EditionDocument115 pagesACT Science Stu 2014 EditionrafiNo ratings yet
- Food Hydrocolloids Volume Issue 2019 (Doi 10.1016 - J.foodhyd.2019.02.023) Huang, Hao Belwal, Tarun Liu, Songbai Duan, Zhenhua Luo, Zis - Novel Multi-Phase Nano-Emulsion Preparation For Co-Loadin PDFDocument37 pagesFood Hydrocolloids Volume Issue 2019 (Doi 10.1016 - J.foodhyd.2019.02.023) Huang, Hao Belwal, Tarun Liu, Songbai Duan, Zhenhua Luo, Zis - Novel Multi-Phase Nano-Emulsion Preparation For Co-Loadin PDFdikativiNo ratings yet
- Extraction Methods, Chemical Characterizations and Biological Activities ofDocument9 pagesExtraction Methods, Chemical Characterizations and Biological Activities ofJUAN C. OVIEDO LOPERANo ratings yet
- Catalase and Hydrogen Peroxide Experiment: Problem: What Happens When A Potato Is Combined With Hydrogen Peroxide?Document1 pageCatalase and Hydrogen Peroxide Experiment: Problem: What Happens When A Potato Is Combined With Hydrogen Peroxide?Big CinemaNo ratings yet
- 1 s2.0 S0141813021025605 MainDocument11 pages1 s2.0 S0141813021025605 MainHanan AqoubNo ratings yet
- Review Article: Fungal Laccases: Production, Function, and Applications in Food ProcessingDocument11 pagesReview Article: Fungal Laccases: Production, Function, and Applications in Food ProcessingRizwan ArshadNo ratings yet
- Savarino Et Al. 2023 - Different Escin TypesDocument19 pagesSavarino Et Al. 2023 - Different Escin TypesBenedikt EngelNo ratings yet
- Ardestani Et Al., 2015Document8 pagesArdestani Et Al., 201515041168No ratings yet
- International Journal of Food Microbiology: ArticleinfoDocument8 pagesInternational Journal of Food Microbiology: ArticleinfoJessica BalganiNo ratings yet
- 1 s2.0 S0144861720302502 MainDocument6 pages1 s2.0 S0144861720302502 Mainعمر البدويNo ratings yet
- Binhayeeding2020 - 1Document12 pagesBinhayeeding2020 - 1Siva ForeviewNo ratings yet
- Lukondeh 2005Document5 pagesLukondeh 2005AleLeonNo ratings yet
- 1-S2.0-S0141813023055484-Main Antioxidant of LP Xingjie WangDocument10 pages1-S2.0-S0141813023055484-Main Antioxidant of LP Xingjie Wangnur rohmanNo ratings yet
- BiokonversiDocument9 pagesBiokonversiAnastasya AmandaNo ratings yet
- (MB) Ortiz-Tena2016 Monosaccharides in CVDocument9 pages(MB) Ortiz-Tena2016 Monosaccharides in CVSacra PsyntergiaNo ratings yet
- Balance de Materiales en Ingenio Central ProgresoDocument11 pagesBalance de Materiales en Ingenio Central ProgresoManuel Jose Tabares MontoyaNo ratings yet
- International Journal of Biological Macromolecules Comparative Study 1-S2.0-S0141813019393699-MainDocument9 pagesInternational Journal of Biological Macromolecules Comparative Study 1-S2.0-S0141813019393699-MainOmar KHELILNo ratings yet
- FullerenosDocument10 pagesFullerenosKaren GomezNo ratings yet
- Foods: Insights Into The Metabolomic Diversity of Latilactobacillus SakeiDocument12 pagesFoods: Insights Into The Metabolomic Diversity of Latilactobacillus SakeiOscar de la ParraNo ratings yet
- Carbohydrate PolymersDocument9 pagesCarbohydrate PolymersAnonymous Wcj4C3jNo ratings yet
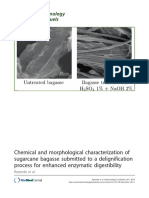
- Chemical and Morphological Characterization of Sugarcane Bagasse Submitted To A Delignification Process For Enhanced Enzymatic DigestibilityDocument19 pagesChemical and Morphological Characterization of Sugarcane Bagasse Submitted To A Delignification Process For Enhanced Enzymatic DigestibilityTabiita Huaman NorabuenaNo ratings yet
- Lactosuero 9 NoDocument24 pagesLactosuero 9 NoMarco Antonio Salcedo HinojosaNo ratings yet
- Anwar Et Al 2008 The Probiotic Lactobacillus Johnsonii NCC 533 Produces High Molecular Mass Inulin From Sucrose byDocument8 pagesAnwar Et Al 2008 The Probiotic Lactobacillus Johnsonii NCC 533 Produces High Molecular Mass Inulin From Sucrose bygugahgiersNo ratings yet
- From By-Product To Valuable Components: Efficient Enzymatic Conversion of Lactose in Whey Using Galactosidase From Streptococcus ThermophilusDocument9 pagesFrom By-Product To Valuable Components: Efficient Enzymatic Conversion of Lactose in Whey Using Galactosidase From Streptococcus ThermophilusAntonela PortaNo ratings yet
- Glycan Processing in Gut MicrobiomesDocument13 pagesGlycan Processing in Gut MicrobiomesinmitamenaNo ratings yet
- Intrinsic Levanase Activity of Bacillus Subtilis 168 Levansucrase (Sacb)Document15 pagesIntrinsic Levanase Activity of Bacillus Subtilis 168 Levansucrase (Sacb)Natalia Norika Yassunaka HataNo ratings yet
- s41598-023-50173-6Document12 pagess41598-023-50173-6dmmbablooNo ratings yet
- Bioaktivni Ugljenihidrati PDFDocument4 pagesBioaktivni Ugljenihidrati PDFmajabulatNo ratings yet
- Ref Khu Mau 1Document9 pagesRef Khu Mau 1HƯƠNG NGUYỄN LÊ NGỌCNo ratings yet
- Levan-Application and PerspectivesDocument17 pagesLevan-Application and PerspectivesDanNo ratings yet
- Foods: Exopolysaccharides Produced by Lactic Acid Bacteria: From Biosynthesis To Health-Promoting PropertiesDocument23 pagesFoods: Exopolysaccharides Produced by Lactic Acid Bacteria: From Biosynthesis To Health-Promoting PropertiesLakshmipathy PNo ratings yet
- MainDocument8 pagesMainsuzuuhNo ratings yet
- Produccion de Acido Lactico A Partir de Suero de LDocument7 pagesProduccion de Acido Lactico A Partir de Suero de LJulieta AhumadaNo ratings yet
- TMP 93 EDocument9 pagesTMP 93 EFrontiersNo ratings yet
- Fermentación BatchDocument8 pagesFermentación BatchJennifer A. PatiñoNo ratings yet
- Accepted Manuscript: Enzyme and Microbial TechnologyDocument41 pagesAccepted Manuscript: Enzyme and Microbial TechnologyTidar MelianiNo ratings yet
- Aerobic Metabolism of Lactic Acid BacteriaDocument12 pagesAerobic Metabolism of Lactic Acid Bacteriaadel strikeNo ratings yet
- The Genus Lactobacillus: A Taxonomic Update: Probiotics and Antimicrobial Proteins December 2012Document11 pagesThe Genus Lactobacillus: A Taxonomic Update: Probiotics and Antimicrobial Proteins December 2012Lisa Purnia CahyaniNo ratings yet
- Complex Coacervation - Encapsulation and Controlled Release of ADocument11 pagesComplex Coacervation - Encapsulation and Controlled Release of AMIRIAM CRISTINA DA SILVA GOMESNo ratings yet
- 10 5923 J Food 20140403 04 PDFDocument6 pages10 5923 J Food 20140403 04 PDFERIKA MARIE BECERRELNo ratings yet
- Degradation of Sugarcane Vinasse in A Full-Scale Methanogenic UASBDocument11 pagesDegradation of Sugarcane Vinasse in A Full-Scale Methanogenic UASBProyectos Plantas De Tratamiento De AguaNo ratings yet
- Enzymatic Synthesis of Carbohydrate-Containing Biomolecules: Advanced ArticleDocument22 pagesEnzymatic Synthesis of Carbohydrate-Containing Biomolecules: Advanced Articleazzaass100% (1)
- Antibacterial Metabolites of Lactic Acid Bacteria: Their Diversity and PropertiesDocument15 pagesAntibacterial Metabolites of Lactic Acid Bacteria: Their Diversity and PropertiesHanaNo ratings yet
- Fermentation 09 00179 v2Document20 pagesFermentation 09 00179 v2ഗീതു സുരേഷ്No ratings yet
- Bernaerts 2017Document12 pagesBernaerts 2017gasparperes922No ratings yet
- Natural Emulsificantes 2Document6 pagesNatural Emulsificantes 2Bruna XavierNo ratings yet
- Lactic Acid Bacteria ProjectDocument17 pagesLactic Acid Bacteria ProjectLaksilu Viduraga PeirisNo ratings yet
- Rumen ADocument11 pagesRumen AnazkasihNo ratings yet
- Yeast and Fermentation Term PaperDocument10 pagesYeast and Fermentation Term PaperSehrish SherryNo ratings yet
- An Overview On Biological Production of Functional Lactose DerivativesDocument9 pagesAn Overview On Biological Production of Functional Lactose DerivativesKukymovNo ratings yet
- Biochemical Characterization and SynergismDocument12 pagesBiochemical Characterization and SynergismJayesh PatidarNo ratings yet
- Food Microbiology: David Laureys, Maarten Aerts, Peter Vandamme, Luc de VuystDocument11 pagesFood Microbiology: David Laureys, Maarten Aerts, Peter Vandamme, Luc de VuystAngeles SuarezNo ratings yet
- Jurnal Lactic Acid-Evan-I1021141064 PDFDocument21 pagesJurnal Lactic Acid-Evan-I1021141064 PDFevanhuangNo ratings yet
- Carbohydrate Polymers: Periaswamy Sivagnanam Saravana, Yeon-Jin Cho, Yong-Beom Park, Hee-Chul Woo, Byung-Soo ChunDocument8 pagesCarbohydrate Polymers: Periaswamy Sivagnanam Saravana, Yeon-Jin Cho, Yong-Beom Park, Hee-Chul Woo, Byung-Soo ChunLeo GarcíaNo ratings yet
- 1 s2.0 S1517838216313740 MainDocument8 pages1 s2.0 S1517838216313740 Main7fkqx7zh7pNo ratings yet
- Pandey Et Al-1999-Biotechnology and Applied Biochemistry PDFDocument13 pagesPandey Et Al-1999-Biotechnology and Applied Biochemistry PDFAlbertochoNo ratings yet
- 1 s2.0 S0308814618308148 MainDocument9 pages1 s2.0 S0308814618308148 MainAusteridad LopezNo ratings yet
- 0 s2.0 S0168165622000268 MainDocument8 pages0 s2.0 S0168165622000268 MainOmar KHELILNo ratings yet
- Potential of Three Probiotic Lactobacilli in Transforming Star Fruit Juice Into Functional BeveragesDocument10 pagesPotential of Three Probiotic Lactobacilli in Transforming Star Fruit Juice Into Functional Beverageskserra.idNo ratings yet
- Biosurfactants From YeastDocument18 pagesBiosurfactants From YeastoniatrdNo ratings yet
- Tabssum Et Al-2018-Journal of Biological EngineeringDocument11 pagesTabssum Et Al-2018-Journal of Biological EngineeringFaisal AwanNo ratings yet
- A Compilation of Ligno-Cellulose Feedstock and Related Research for Feed, Food and EnergyFrom EverandA Compilation of Ligno-Cellulose Feedstock and Related Research for Feed, Food and EnergyNo ratings yet
- Beetles Infesting Woolens RreztqDocument3 pagesBeetles Infesting Woolens RreztqsmurfsiftNo ratings yet
- Pit and Fissure SealantsDocument35 pagesPit and Fissure SealantsshrutiNo ratings yet
- Pengaruh Konsentrasi Carboxy Metil Cellulose (CMC) Dan Gliserol Terhadap Karakteristik Edible Film BEKATUL PADI (Oryza Sativa)Document10 pagesPengaruh Konsentrasi Carboxy Metil Cellulose (CMC) Dan Gliserol Terhadap Karakteristik Edible Film BEKATUL PADI (Oryza Sativa)UjicobaNo ratings yet
- Hazardous Area Classification (HAC) Using QuadventDocument10 pagesHazardous Area Classification (HAC) Using QuadventRiad Riad100% (1)
- Indian Standard: Bamboo Mat Corrugated Sheets - SpecificationDocument15 pagesIndian Standard: Bamboo Mat Corrugated Sheets - SpecificationJunaidAhmedNo ratings yet
- LaMotte 3674-01 DC1200-FL Fluoride Colorimeter Kit InstructionsDocument4 pagesLaMotte 3674-01 DC1200-FL Fluoride Colorimeter Kit InstructionsPromagEnviro.comNo ratings yet
- Organic Chemistry NotesDocument21 pagesOrganic Chemistry NotesBobbyWhiteNo ratings yet
- Ch19 Wade ChemistryDocument58 pagesCh19 Wade ChemistrySunnyd1013No ratings yet
- Daftar PustakaDocument3 pagesDaftar PustakaMis SakqNo ratings yet
- PressedDocument92 pagesPressedSimo AfamaswaagerNo ratings yet
- Aspartate Aminotransferase (Ast/Got)Document2 pagesAspartate Aminotransferase (Ast/Got)Ivana BajunovicNo ratings yet
- 12 Volt 10 Ampere Solar Charge Controller ManualDocument26 pages12 Volt 10 Ampere Solar Charge Controller ManualONETRUEGEMLLC50% (2)
- Atoms, Isotopes, Ions, and MoleculesDocument67 pagesAtoms, Isotopes, Ions, and MoleculesRochelle NuguidNo ratings yet
- Part-1 For FFDocument73 pagesPart-1 For FFvictorhugodiaz20085603No ratings yet
- Anode InfoDocument5 pagesAnode InfoEberg NlnoNo ratings yet
- Detection of Pyrrolizidine Alkaloids in A Few Medicinal PlantsDocument9 pagesDetection of Pyrrolizidine Alkaloids in A Few Medicinal PlantsfabriziozaraNo ratings yet
- Introduction To Soil Mechanics: Level-1Document6 pagesIntroduction To Soil Mechanics: Level-1pratik gamaraNo ratings yet
- Presented To: Dr. Islam Ullah Khan Presented By: Khadeeja Ikram Roll No: 0164-BH-CHEM-11Document45 pagesPresented To: Dr. Islam Ullah Khan Presented By: Khadeeja Ikram Roll No: 0164-BH-CHEM-11Abiyot NeguNo ratings yet
- Aapfco Product Label Guide: Association of American Plant Food Control Officials 2019Document31 pagesAapfco Product Label Guide: Association of American Plant Food Control Officials 2019ronalit malintadNo ratings yet
- 552 Shazia (Rubber)Document22 pages552 Shazia (Rubber)Anishah ChaudheryNo ratings yet
- Unit 3 Pressure GaugeDocument11 pagesUnit 3 Pressure GaugeUv sai likithNo ratings yet
- Photoelectrictransducer 131228100036 Phpapp02Document15 pagesPhotoelectrictransducer 131228100036 Phpapp02jishnuNo ratings yet
- G7-Biology Second Term 2078Document21 pagesG7-Biology Second Term 2078Skeltten MinecraftNo ratings yet
- The Difference Between Cellulose and ChitinDocument6 pagesThe Difference Between Cellulose and Chitincindy khairunnisaNo ratings yet
- Short Notes by SK SirDocument8 pagesShort Notes by SK SirJay MeenaNo ratings yet
- MT Powder MetallurgyDocument74 pagesMT Powder MetallurgyAhad RazzNo ratings yet
- Ashish Kumar Patel (Assistant Professor) Mechanical Engineering Department Lovely Professional UniversityDocument16 pagesAshish Kumar Patel (Assistant Professor) Mechanical Engineering Department Lovely Professional UniversitySachin KumarNo ratings yet
- BS en 1514 1 1997Document22 pagesBS en 1514 1 1997Phaneendra Tirumani100% (2)