Download as pdf or txt
You might also like
- Free Download Nutrition Concepts and Controversies 14Th Edition Frances Sizer Full Chapter PDFDocument51 pagesFree Download Nutrition Concepts and Controversies 14Th Edition Frances Sizer Full Chapter PDFpatrica.ramirez337100% (22)
- Nickel Powerhouse CDocument49 pagesNickel Powerhouse CgussalimNo ratings yet
- Preview of Nutrition Through The Life Cycle 3rd EditionDocument20 pagesPreview of Nutrition Through The Life Cycle 3rd EditionHAFIZ MUHAMMAD IMTIAZ100% (1)
- Dietary Reference Intakes (DRI)Document1 pageDietary Reference Intakes (DRI)ninik maathiaNo ratings yet
- ebook download Nutrition: concepts and controversies Fourteenth Edition Webb - eBook PDF all chapterDocument54 pagesebook download Nutrition: concepts and controversies Fourteenth Edition Webb - eBook PDF all chaptermuxaankiener100% (2)
- Ebook Nutrition Concepts and Controversies PDF Full Chapter PDFDocument67 pagesEbook Nutrition Concepts and Controversies PDF Full Chapter PDFmegan.king288100% (35)
- PDF Nutrition Concepts and Controversies Fourteenth Edition Webb Ebook Full ChapterDocument53 pagesPDF Nutrition Concepts and Controversies Fourteenth Edition Webb Ebook Full Chapterashley.roberts341100% (2)
- Textbook Nutrition Concepts and Controversies 14Th Edition Frances Sizer Ebook All Chapter PDFDocument53 pagesTextbook Nutrition Concepts and Controversies 14Th Edition Frances Sizer Ebook All Chapter PDFkaren.eakin573100% (15)
- ebook download Nutrition Concepts and Controversies : 14th Edition Frances Sizer - eBook PDF all chapterDocument54 pagesebook download Nutrition Concepts and Controversies : 14th Edition Frances Sizer - eBook PDF all chapterberjeblatz100% (4)
- Nutrition Concepts and Controversies Fourteenth Edition Webb Full ChapterDocument67 pagesNutrition Concepts and Controversies Fourteenth Edition Webb Full Chapterjeff.tinker162100% (9)
- Krishna Final PresentationDocument29 pagesKrishna Final Presentationyanshutradelink.pvtNo ratings yet
- Full download Nutrition: concepts and controversies Fourteenth Edition Webb file pdf all chapter on 2024Document44 pagesFull download Nutrition: concepts and controversies Fourteenth Edition Webb file pdf all chapter on 2024koricbondiaNo ratings yet
- (Download PDF) Nutrition Concepts and Controversies Fourteenth Edition Webb Full Chapter PDFDocument69 pages(Download PDF) Nutrition Concepts and Controversies Fourteenth Edition Webb Full Chapter PDFcokuzajokke22100% (8)
- Coral ReefsDocument4 pagesCoral ReefsFriida BCNo ratings yet
- Wastewater Treatment Plant Privatization IDocument45 pagesWastewater Treatment Plant Privatization Iashraf.rahim139No ratings yet
- Assignment Environmental Science IBDocument3 pagesAssignment Environmental Science IBCosmicBlue24No ratings yet
- QATAR6Document4 pagesQATAR6Cássio MagalhãesNo ratings yet
- Contingency TablesDocument5 pagesContingency TablesMMTNo ratings yet
- 2011 ... Safa, Biosorption of Direct Red-31 and DODocument15 pages2011 ... Safa, Biosorption of Direct Red-31 and DOarturo_morinNo ratings yet
- Second Periodical Test in Science 5Document6 pagesSecond Periodical Test in Science 5raquel alvarezNo ratings yet
- L02 - The Philippine Coastal Zone - An Endangered EnvironmenDocument25 pagesL02 - The Philippine Coastal Zone - An Endangered EnvironmenehmNo ratings yet
- Wcms 232060Document46 pagesWcms 232060madeusmoNo ratings yet
- 2016 Code PDocument43 pages2016 Code PAsif AnsariNo ratings yet
- DIference Betwen Rivers and LakeDocument2 pagesDIference Betwen Rivers and LakeAngelo ThiagoNo ratings yet
- Epidemiology of Malaria, Clinical Features, Treatment and Its ControlDocument157 pagesEpidemiology of Malaria, Clinical Features, Treatment and Its ControlJ Nuchin100% (2)
- CEM 3005W Typed Notes Reaction Classes and Pericyclic Reactions 2013Document12 pagesCEM 3005W Typed Notes Reaction Classes and Pericyclic Reactions 2013Zama MakhathiniNo ratings yet
- Procesamiento de Información EstadísticaDocument8 pagesProcesamiento de Información EstadísticaDanna Evioleth Torres RamosNo ratings yet
- HTML To PDFDocument1 pageHTML To PDFYesenia GaleoteNo ratings yet
- Concept of Secondary ProductivityDocument16 pagesConcept of Secondary ProductivitylithunNo ratings yet
- Word AsistensiDocument2 pagesWord AsistensikegitankuliahbisaNo ratings yet
- Navg Annual 2005Document131 pagesNavg Annual 2005Radim StuchlikNo ratings yet
- 2012 Dse MSDocument14 pages2012 Dse MShq5sy9gjb5No ratings yet
- BEVAE-181Document20 pagesBEVAE-181shaliniarora19704No ratings yet
- Remotesensing 14 04868Document29 pagesRemotesensing 14 04868WIta DeuterNo ratings yet
- Primary Production Required To Sustain Global FisheriesDocument5 pagesPrimary Production Required To Sustain Global Fisheries123456No ratings yet
- CAPE Env. Science 2012 U1 P2Document9 pagesCAPE Env. Science 2012 U1 P2Christina FrancisNo ratings yet
- Aakash ANTHE Junior Ques Class 9 2015 PDFDocument8 pagesAakash ANTHE Junior Ques Class 9 2015 PDFhamidNo ratings yet
- Microbiology Article Membrane RecoveriesDocument10 pagesMicrobiology Article Membrane RecoveriesSofia CbaNo ratings yet
- Solved MCQsDocument5 pagesSolved MCQsAli Raza BalochNo ratings yet
- Maddipadu Anc 2021-22Document43 pagesMaddipadu Anc 2021-22Amrutha Heart HospitalNo ratings yet
- 28. Dec 2023 BEVAE-181 IGNOUAssignmentGuru.comDocument20 pages28. Dec 2023 BEVAE-181 IGNOUAssignmentGuru.comsubhampaul609No ratings yet
- QuestionDocument11 pagesQuestionkembara08No ratings yet
- Agarwal OCP Rainwater 1986Document10 pagesAgarwal OCP Rainwater 1986Popescu DragosNo ratings yet
- EnvScience 2014 U1 P2Document10 pagesEnvScience 2014 U1 P2leahblanc05No ratings yet
- Environmental Science DraftDocument53 pagesEnvironmental Science DraftMouricia JonesNo ratings yet
- Bandhan Focused Equity FundDocument10 pagesBandhan Focused Equity FundArmstrong CapitalNo ratings yet
- TC La Ss Es: LRDI - 01Document24 pagesTC La Ss Es: LRDI - 01Ankita GuptaNo ratings yet
- Seco 2020Document39 pagesSeco 2020yosyNo ratings yet
- AIPGE Dental Paper of MDSDocument34 pagesAIPGE Dental Paper of MDSMohit ChandraNo ratings yet
- ALL About Sta. Praxedes and Its MunicipalityDocument5 pagesALL About Sta. Praxedes and Its MunicipalityJeric CrashByte LedamaNo ratings yet
- 2012 Dse Bio 1 MS 1Document6 pages2012 Dse Bio 1 MS 1peakNo ratings yet
- IB Ecology Review Key 2019Document8 pagesIB Ecology Review Key 2019Yasmine Ouled Kaddour El HallaouiNo ratings yet
- Owolabi 1994Document6 pagesOwolabi 1994AbassyacoubouNo ratings yet
- Adobe Scan 6 Feb 2024Document2 pagesAdobe Scan 6 Feb 2024kohinoorcheema06No ratings yet
- CEA HomeDocument33 pagesCEA HomeYmon TuallaNo ratings yet
- Proper Use of Capillary Number in Chemical FloodingDocument12 pagesProper Use of Capillary Number in Chemical FloodingHu GuoNo ratings yet
- Introduction To Academic WritingDocument232 pagesIntroduction To Academic WritingMahfoodh Aljuboury100% (3)
- Biodiversity DriversDocument15 pagesBiodiversity DriversAbhinav UpadhyayNo ratings yet
- BEVAE 181 June2010 June2023 IGNOUAssignmentGuru.comDocument120 pagesBEVAE 181 June2010 June2023 IGNOUAssignmentGuru.compratikdnbfamNo ratings yet
- Specific Paper: Question BookletDocument32 pagesSpecific Paper: Question BookletrpsirNo ratings yet
- GenderStereotyping VAW ReportDocument69 pagesGenderStereotyping VAW ReportAbhinavSinghNo ratings yet
- Challenge of Maan - Challenge of Maan - JansattaDocument4 pagesChallenge of Maan - Challenge of Maan - JansattaAbhinavSinghNo ratings yet
- Juvenile JusticeDocument56 pagesJuvenile JusticeAbhinavSinghNo ratings yet
- 76.drug Market - Drug Market - JansattaDocument3 pages76.drug Market - Drug Market - JansattaAbhinavSinghNo ratings yet
- Scra ActDocument43 pagesScra ActAbhinavSinghNo ratings yet
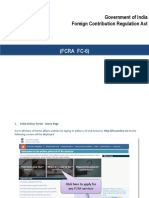
- (FCRA FC-6) : Government of India Foreign Contribution Regulation ActDocument21 pages(FCRA FC-6) : Government of India Foreign Contribution Regulation ActAbhinavSinghNo ratings yet
- Chapter-IX Account of CompaniesDocument47 pagesChapter-IX Account of CompaniesAbhinavSinghNo ratings yet
- Dgca PDFDocument37 pagesDgca PDFAbhinavSingh100% (1)
- IcaDocument6 pagesIcaAbhinavSinghNo ratings yet
- SEBI Takeover CodeDocument5 pagesSEBI Takeover CodeAbhinavSinghNo ratings yet
- Monkeys and ForestsDocument19 pagesMonkeys and ForestsBhupathi SukeshNo ratings yet
- IUCN Red List of Threatened SpeciesDocument12 pagesIUCN Red List of Threatened SpeciesssaygunNo ratings yet
- Ciclidos 6-1Document859 pagesCiclidos 6-1Franklin RadaNo ratings yet
- MPC 15Document63 pagesMPC 15IcNo ratings yet
- South Asian Amphibians - Taxonomy, Diversity and Conservation StatusDocument15 pagesSouth Asian Amphibians - Taxonomy, Diversity and Conservation StatusRegina Joyce E. FerrerNo ratings yet
- PterDocument40 pagesPterSahaya AnthonyNo ratings yet
- 10.2305 IUCN - UK.1998.RLTS.T38596A10131047.en PDFDocument5 pages10.2305 IUCN - UK.1998.RLTS.T38596A10131047.en PDFpujimentariNo ratings yet
- Biodiversity Conservation: Module - 5Document22 pagesBiodiversity Conservation: Module - 5Pratyush Goel100% (1)
- Conservation-Dependent Species - WikipediaDocument3 pagesConservation-Dependent Species - WikipediadearbhupiNo ratings yet
- MP National ParkDocument9 pagesMP National Parkankita100% (1)
- Biodiversity Analysis Update For Georgia 1Document89 pagesBiodiversity Analysis Update For Georgia 1opentaps0% (1)
- PK Article 24380 en 1 PDFDocument341 pagesPK Article 24380 en 1 PDFKseniia MaslakovaNo ratings yet
- Supplementary Materials For: Protected Areas and Effective Biodiversity ConservationDocument29 pagesSupplementary Materials For: Protected Areas and Effective Biodiversity ConservationCássio Batista MarconNo ratings yet
- End-Of-Term Test 2Document17 pagesEnd-Of-Term Test 2Le Cao DuyNo ratings yet
- The International Union For The Conservation of Nature (IUCN) Is The World's Main Authority On TheDocument2 pagesThe International Union For The Conservation of Nature (IUCN) Is The World's Main Authority On Theprantik32No ratings yet
- National Parks 1Document93 pagesNational Parks 1Manoj GoyalNo ratings yet
- ENVIRONMENTAL ETHICS ANTHROPOCENTRIC TO ECO-CENTRIC APPROACH - Harsha NDocument10 pagesENVIRONMENTAL ETHICS ANTHROPOCENTRIC TO ECO-CENTRIC APPROACH - Harsha Nmahendra prajapatiNo ratings yet
- Field Guidebook On The Native Trees Within The Quirino Forest LandscapeDocument134 pagesField Guidebook On The Native Trees Within The Quirino Forest LandscapeSteve TanNo ratings yet
- Sample PDF of STD 11 and 12 Based MHT Cet Triumph Biology Maharashtra State BoardDocument19 pagesSample PDF of STD 11 and 12 Based MHT Cet Triumph Biology Maharashtra State Boardvrushalin40No ratings yet
- 10.2305 IUCN - UK.2019-3.RLTS.T61911798A61911825.enDocument9 pages10.2305 IUCN - UK.2019-3.RLTS.T61911798A61911825.enMeche KakaNo ratings yet
- DIVERSITY: A Compilation of Organisms Exhibited at National Museum of Natural HistoryDocument106 pagesDIVERSITY: A Compilation of Organisms Exhibited at National Museum of Natural HistoryDamNo ratings yet
- Environment PPT Template 022Document20 pagesEnvironment PPT Template 022Kalai ShanNo ratings yet
- Red Data BookDocument2 pagesRed Data BookHamza shehrozNo ratings yet
- 100 Questions On EnvironmentDocument20 pages100 Questions On EnvironmentvgasNo ratings yet
- RL 549.3 003 v.4Document337 pagesRL 549.3 003 v.4Hazi TelapokaNo ratings yet
- On The Halophytic Nature of MangrovesDocument6 pagesOn The Halophytic Nature of Mangrovestanjidur rahmanNo ratings yet
- Lakhmir Singh Solutions For Class 8 Science Chapter 7Document12 pagesLakhmir Singh Solutions For Class 8 Science Chapter 7Purnabi JenaNo ratings yet
- Graduate Student Work: A Model For Direct and Mutually Beneficial Community-University PartnershipsDocument1 pageGraduate Student Work: A Model For Direct and Mutually Beneficial Community-University Partnershipsapi-253296477No ratings yet
- 14491-Article Text-1086091-1-10-20230701 (Inggris)Document13 pages14491-Article Text-1086091-1-10-20230701 (Inggris)albian mubarakNo ratings yet