
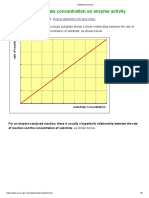
Competitive Inhibition
Competitive Inhibition
You might also like
- Chemo Chart Final 121509Document7 pagesChemo Chart Final 121509José Mauricio Peñaloza100% (4)
- Enzyme Part 1-5 Vikneswaran 260110132004Document7 pagesEnzyme Part 1-5 Vikneswaran 260110132004Vikneswaran VîçkýNo ratings yet
- Enzyme InhibitionDocument4 pagesEnzyme InhibitionNisha ManasNo ratings yet
- Past Year CreDocument22 pagesPast Year CreNURUL SYAHIRAH BINTI ABDUL HALIMNo ratings yet
- Bio Chapter 3Document103 pagesBio Chapter 3Areej AlissaNo ratings yet
- Lecture 15: Enzyme Inhibition: Biochemistry I Fall Term, 2003Document4 pagesLecture 15: Enzyme Inhibition: Biochemistry I Fall Term, 2003archerofthestarsNo ratings yet
- Lectures - Biochemistry 1 - 2021-2022 Prof Version 6.01 - 23-24-231-246Document16 pagesLectures - Biochemistry 1 - 2021-2022 Prof Version 6.01 - 23-24-231-246Ken M'voulaboloNo ratings yet
- Enzyme KineticsDocument8 pagesEnzyme KineticsronojoysenguptaNo ratings yet
- 04 - Enzyme Inhibition PDFDocument34 pages04 - Enzyme Inhibition PDFJavier Torres100% (1)
- Interactions Between Antigen and AntibodyDocument25 pagesInteractions Between Antigen and AntibodyvmshanesNo ratings yet
- Enzyme-Kinetics and Specificity: Modified by Chung-Ming Lin Department of Biotechnology Ming Chuan UniversityDocument58 pagesEnzyme-Kinetics and Specificity: Modified by Chung-Ming Lin Department of Biotechnology Ming Chuan University陳家琪No ratings yet
- A Graphical Method For Determining Inhibition ConstantsDocument4 pagesA Graphical Method For Determining Inhibition ConstantsPriyanshu Priyadarshi MeherNo ratings yet
- Estimación Parámetros DoranDocument4 pagesEstimación Parámetros DoranfernandaNo ratings yet
- Michaelis Menten EquationDocument4 pagesMichaelis Menten Equationsumant5shringari100% (1)
- Types of Inhibirtion GreatDocument3 pagesTypes of Inhibirtion Greatteam TSOTARENo ratings yet
- Enzyme KineticsDocument13 pagesEnzyme KineticsKhushbu JainNo ratings yet
- Enzyme KineticsDocument10 pagesEnzyme KineticsQuenneBelocuraNo ratings yet
- EnzomologyDocument26 pagesEnzomologyToga Brandon100% (1)
- Optimal Placement and Sizing of Static VDocument5 pagesOptimal Placement and Sizing of Static VDa VidNo ratings yet
- Enzymology (GEB-2101) Lecture 3Document15 pagesEnzymology (GEB-2101) Lecture 3Niloy GhoshNo ratings yet
- BCM Enzyme II 200lDocument46 pagesBCM Enzyme II 200lewoozino1234No ratings yet
- Answer: Enzyme Kinetics (Michaelis Menten/Lineweaver Burke/Eadie-Hostee)Document2 pagesAnswer: Enzyme Kinetics (Michaelis Menten/Lineweaver Burke/Eadie-Hostee)Ryan Borazon GarciaNo ratings yet
- Characteristics of The Non-Sinusoidal Coefficient of The Voltage at Node Containing A Thyristor Compensator in Presence of HarmonicsDocument4 pagesCharacteristics of The Non-Sinusoidal Coefficient of The Voltage at Node Containing A Thyristor Compensator in Presence of HarmonicsZellagui EnergyNo ratings yet
- Voltage Stability Analysis BY Simulated Annealing AlgorithmDocument7 pagesVoltage Stability Analysis BY Simulated Annealing AlgorithmInternational Journal of Engineering and TechniquesNo ratings yet
- Question CH06+answer PDFDocument8 pagesQuestion CH06+answer PDFCris-Anne Juangco III100% (1)
- 5.2 - Enzyme Parameters - Chemistry LibreTextsDocument1 page5.2 - Enzyme Parameters - Chemistry LibreTextsteam TSOTARENo ratings yet
- BioOrganic Notes Part-4 FinalDocument16 pagesBioOrganic Notes Part-4 FinalAnupama GuptaNo ratings yet
- Simple Kinetics of Enzyme ActionDocument7 pagesSimple Kinetics of Enzyme ActionRavindra Mani TiwariNo ratings yet
- Paper 2Document7 pagesPaper 2Ruben Barros GodoyNo ratings yet
- Antigen Antibody ReactionDocument16 pagesAntigen Antibody ReactionNadzierah RazakNo ratings yet
- Relationship Between Ki and IC50Document10 pagesRelationship Between Ki and IC50Jacqueto ZephyrNo ratings yet
- Thevenin's Theorem and Maximum Power TransferDocument26 pagesThevenin's Theorem and Maximum Power TransferNathaniel PanganibanNo ratings yet
- Lecture-19 Chapter 3Document14 pagesLecture-19 Chapter 3ktrone88No ratings yet
- EnzymeKinetics by P.C. Misra Professor, Department of Biochemistry Lucknow University, Lucknow-226 007Document21 pagesEnzymeKinetics by P.C. Misra Professor, Department of Biochemistry Lucknow University, Lucknow-226 007Dr. SHIVA AITHALNo ratings yet
- Non-Linear Pharmacokinetics: Biopharmaceutics (Ashik Sir)Document9 pagesNon-Linear Pharmacokinetics: Biopharmaceutics (Ashik Sir)Hasan Al MasudNo ratings yet
- GSCL Diferences Kinetic EnzymeDocument3 pagesGSCL Diferences Kinetic Enzymejoenni hansNo ratings yet
- 9.07 Theories of Reaction Rates PDFDocument6 pages9.07 Theories of Reaction Rates PDFAbdelfattah Mohamed OufNo ratings yet
- Biochem ConceptsDocument9 pagesBiochem Conceptsdanena88No ratings yet
- Exam II ReviewDocument9 pagesExam II ReviewShira ChoiNo ratings yet
- Mechaelis-Menten KineticsDocument7 pagesMechaelis-Menten KineticsfayeNo ratings yet
- The Molecular Symmetry GroupDocument24 pagesThe Molecular Symmetry GroupWippetsxzNo ratings yet
- Chapter 6 Parallel DC CircuitsDocument72 pagesChapter 6 Parallel DC CircuitsAbdullrahman Al-ShammaaNo ratings yet
- Mikales Menten Great With Green Book AlsoDocument8 pagesMikales Menten Great With Green Book AlsoMOSES MILLERNo ratings yet
- Facts and PQ Unit-IIIDocument9 pagesFacts and PQ Unit-IIIAniket ANo ratings yet
- A Steverson Chapter 11 Pages 416-468Document54 pagesA Steverson Chapter 11 Pages 416-468Gerson NetoNo ratings yet
- Fundamentals of Electrical Circuits AnalysisDocument8 pagesFundamentals of Electrical Circuits AnalysisArcturus VoidNo ratings yet
- Effect of Battery Energy Storage System On Load Frequency Control Considering Governor Deadband and Generation Rate ConstraintDocument7 pagesEffect of Battery Energy Storage System On Load Frequency Control Considering Governor Deadband and Generation Rate ConstraintzahidNo ratings yet
- Ch06 QuizDocument5 pagesCh06 QuizcatlovesfoodNo ratings yet
- LAB 3 - NewDocument5 pagesLAB 3 - Newzaaa asdadsasdaNo ratings yet
- Kuzmic KobsTight 20220825 001Document7 pagesKuzmic KobsTight 20220825 001Petr KuzmicNo ratings yet
- Enzyme Kinetics: CHEM 3501 The University of Memphis Foundations of Bioorganic Chemistry Lab Department of ChemistryDocument6 pagesEnzyme Kinetics: CHEM 3501 The University of Memphis Foundations of Bioorganic Chemistry Lab Department of ChemistryMohammad AliNo ratings yet
- Midnight HWDocument3 pagesMidnight HWDont MeNo ratings yet
- Atkins EnzimasDocument5 pagesAtkins EnzimasConstanza Espinoza LaraNo ratings yet
- Derivations OF: Enzyme Kinetics (I)Document20 pagesDerivations OF: Enzyme Kinetics (I)Sabari Krishnan B B100% (1)
- Enzmology RevisionDocument8 pagesEnzmology RevisionRyan Fortune AludaNo ratings yet
- Lab 10 PDFDocument3 pagesLab 10 PDFFaiyaz ShaikhNo ratings yet
- Hypervariable Regions: Nature of Antigen-Antibody Reactions Lock and Key ConceptDocument7 pagesHypervariable Regions: Nature of Antigen-Antibody Reactions Lock and Key ConceptVenkata Ramana Murthy VasupilliNo ratings yet
- Capacitor Placement in Unbalanced Distribution Networks To Minimize Reactive and Unbalance Power LossesDocument8 pagesCapacitor Placement in Unbalanced Distribution Networks To Minimize Reactive and Unbalance Power LossesJones LennonNo ratings yet
- Chapter11-Two Ways AnovaDocument26 pagesChapter11-Two Ways AnovaRaluca NegruNo ratings yet
- Reviews in Computational ChemistryFrom EverandReviews in Computational ChemistryAbby L. ParrillNo ratings yet
- On The Integration of in Silico Drug Design Methods For Drug RepurposingDocument7 pagesOn The Integration of in Silico Drug Design Methods For Drug RepurposingdarmaNo ratings yet
- Indian Company ListDocument256 pagesIndian Company ListRaj SaNo ratings yet
- Mencari Selisih (1) EDITDocument426 pagesMencari Selisih (1) EDITlukiharjantiNo ratings yet
- Journal June 2019Document36 pagesJournal June 2019rammvr05No ratings yet
- Nursing Pharmacology BSN2Document9 pagesNursing Pharmacology BSN2Ria Angely PanagaNo ratings yet
- Presentation Adverse Drug ReactionDocument16 pagesPresentation Adverse Drug ReactionAG STXLEZNo ratings yet
- Anda - of AndaDocument46 pagesAnda - of AndaTarikNo ratings yet
- Question Paper DSSSB Phamacist April 2015Document29 pagesQuestion Paper DSSSB Phamacist April 2015Sahil KumarNo ratings yet
- BCS ClassDocument8 pagesBCS Classrina wijayantiNo ratings yet
- Handout On Prescription WritingDocument10 pagesHandout On Prescription WritingAbdiNo ratings yet
- Biopharmaceutics Supplemental NotesDocument19 pagesBiopharmaceutics Supplemental NotesChristine Joy De Ocampo0% (1)
- Inactive Ingredient Search For Approved Dhtghrug ProductsDocument2 pagesInactive Ingredient Search For Approved Dhtghrug ProductsDang Chi CongNo ratings yet
- Daftar Pustaka SterilDocument3 pagesDaftar Pustaka SterilNurhayati Nasrid BobaNo ratings yet
- Marketing ExamDocument42 pagesMarketing ExamMarianne CruzNo ratings yet
- HNMC CV Formulary PDFDocument129 pagesHNMC CV Formulary PDFAnonymous F1xtU2RNo ratings yet
- CTD Dossier Preparation: K. Srikantha Reddy K. Srikantha ReddyDocument46 pagesCTD Dossier Preparation: K. Srikantha Reddy K. Srikantha Reddyfadli100% (1)
- Nfi 2011Document804 pagesNfi 2011indmale_0070% (1)
- Anticancer DrugsDocument6 pagesAnticancer DrugsmidhunNo ratings yet
- Receptor Profile of Selected Anti-Psychotic Drugs: Lovely Liana Clapis Sittie Rahani CanacanDocument42 pagesReceptor Profile of Selected Anti-Psychotic Drugs: Lovely Liana Clapis Sittie Rahani CanacanKevin ChanNo ratings yet
- 2 TIVA - TCI RevisionDocument25 pages2 TIVA - TCI RevisionMora GalliNo ratings yet
- Drug Development and Regulatory Abbreviation SheetDocument16 pagesDrug Development and Regulatory Abbreviation SheetAdamNo ratings yet
- Hetero Corp Presentation 2016Document23 pagesHetero Corp Presentation 2016Evan TanswariNo ratings yet
- Cefuroxime 1st PDF TRIAL PDFDocument1 pageCefuroxime 1st PDF TRIAL PDFRyanNo ratings yet
- Daftar Inventaris Buku FFDocument71 pagesDaftar Inventaris Buku FFcakra gilingNo ratings yet
- Decreased Warfarin Effect After Initiation of High-Protein, Low-Carbohydrate DietsDocument5 pagesDecreased Warfarin Effect After Initiation of High-Protein, Low-Carbohydrate DietsDinaNo ratings yet
- Laporan DakDocument168 pagesLaporan DaknurhasanahNo ratings yet
- ACTD Administrative DataDocument14 pagesACTD Administrative DataWilliam ChandraNo ratings yet
- Orally Disintegrating Tablets: A Novel Approach For MedicationDocument8 pagesOrally Disintegrating Tablets: A Novel Approach For MedicationyNo ratings yet
- Pharmaceutical Care vs. Clinical PharmacyDocument2 pagesPharmaceutical Care vs. Clinical PharmacyKaren Park100% (1)
























































































Download as docx, pdf, or txt
You might also like
- Chemo Chart Final 121509Document7 pagesChemo Chart Final 121509José Mauricio Peñaloza100% (4)
- Enzyme Part 1-5 Vikneswaran 260110132004Document7 pagesEnzyme Part 1-5 Vikneswaran 260110132004Vikneswaran VîçkýNo ratings yet
- Enzyme InhibitionDocument4 pagesEnzyme InhibitionNisha ManasNo ratings yet
- Past Year CreDocument22 pagesPast Year CreNURUL SYAHIRAH BINTI ABDUL HALIMNo ratings yet
- Bio Chapter 3Document103 pagesBio Chapter 3Areej AlissaNo ratings yet
- Lecture 15: Enzyme Inhibition: Biochemistry I Fall Term, 2003Document4 pagesLecture 15: Enzyme Inhibition: Biochemistry I Fall Term, 2003archerofthestarsNo ratings yet
- Lectures - Biochemistry 1 - 2021-2022 Prof Version 6.01 - 23-24-231-246Document16 pagesLectures - Biochemistry 1 - 2021-2022 Prof Version 6.01 - 23-24-231-246Ken M'voulaboloNo ratings yet
- Enzyme KineticsDocument8 pagesEnzyme KineticsronojoysenguptaNo ratings yet
- 04 - Enzyme Inhibition PDFDocument34 pages04 - Enzyme Inhibition PDFJavier Torres100% (1)
- Interactions Between Antigen and AntibodyDocument25 pagesInteractions Between Antigen and AntibodyvmshanesNo ratings yet
- Enzyme-Kinetics and Specificity: Modified by Chung-Ming Lin Department of Biotechnology Ming Chuan UniversityDocument58 pagesEnzyme-Kinetics and Specificity: Modified by Chung-Ming Lin Department of Biotechnology Ming Chuan University陳家琪No ratings yet
- A Graphical Method For Determining Inhibition ConstantsDocument4 pagesA Graphical Method For Determining Inhibition ConstantsPriyanshu Priyadarshi MeherNo ratings yet
- Estimación Parámetros DoranDocument4 pagesEstimación Parámetros DoranfernandaNo ratings yet
- Michaelis Menten EquationDocument4 pagesMichaelis Menten Equationsumant5shringari100% (1)
- Types of Inhibirtion GreatDocument3 pagesTypes of Inhibirtion Greatteam TSOTARENo ratings yet
- Enzyme KineticsDocument13 pagesEnzyme KineticsKhushbu JainNo ratings yet
- Enzyme KineticsDocument10 pagesEnzyme KineticsQuenneBelocuraNo ratings yet
- EnzomologyDocument26 pagesEnzomologyToga Brandon100% (1)
- Optimal Placement and Sizing of Static VDocument5 pagesOptimal Placement and Sizing of Static VDa VidNo ratings yet
- Enzymology (GEB-2101) Lecture 3Document15 pagesEnzymology (GEB-2101) Lecture 3Niloy GhoshNo ratings yet
- BCM Enzyme II 200lDocument46 pagesBCM Enzyme II 200lewoozino1234No ratings yet
- Answer: Enzyme Kinetics (Michaelis Menten/Lineweaver Burke/Eadie-Hostee)Document2 pagesAnswer: Enzyme Kinetics (Michaelis Menten/Lineweaver Burke/Eadie-Hostee)Ryan Borazon GarciaNo ratings yet
- Characteristics of The Non-Sinusoidal Coefficient of The Voltage at Node Containing A Thyristor Compensator in Presence of HarmonicsDocument4 pagesCharacteristics of The Non-Sinusoidal Coefficient of The Voltage at Node Containing A Thyristor Compensator in Presence of HarmonicsZellagui EnergyNo ratings yet
- Voltage Stability Analysis BY Simulated Annealing AlgorithmDocument7 pagesVoltage Stability Analysis BY Simulated Annealing AlgorithmInternational Journal of Engineering and TechniquesNo ratings yet
- Question CH06+answer PDFDocument8 pagesQuestion CH06+answer PDFCris-Anne Juangco III100% (1)
- 5.2 - Enzyme Parameters - Chemistry LibreTextsDocument1 page5.2 - Enzyme Parameters - Chemistry LibreTextsteam TSOTARENo ratings yet
- BioOrganic Notes Part-4 FinalDocument16 pagesBioOrganic Notes Part-4 FinalAnupama GuptaNo ratings yet
- Simple Kinetics of Enzyme ActionDocument7 pagesSimple Kinetics of Enzyme ActionRavindra Mani TiwariNo ratings yet
- Paper 2Document7 pagesPaper 2Ruben Barros GodoyNo ratings yet
- Antigen Antibody ReactionDocument16 pagesAntigen Antibody ReactionNadzierah RazakNo ratings yet
- Relationship Between Ki and IC50Document10 pagesRelationship Between Ki and IC50Jacqueto ZephyrNo ratings yet
- Thevenin's Theorem and Maximum Power TransferDocument26 pagesThevenin's Theorem and Maximum Power TransferNathaniel PanganibanNo ratings yet
- Lecture-19 Chapter 3Document14 pagesLecture-19 Chapter 3ktrone88No ratings yet
- EnzymeKinetics by P.C. Misra Professor, Department of Biochemistry Lucknow University, Lucknow-226 007Document21 pagesEnzymeKinetics by P.C. Misra Professor, Department of Biochemistry Lucknow University, Lucknow-226 007Dr. SHIVA AITHALNo ratings yet
- Non-Linear Pharmacokinetics: Biopharmaceutics (Ashik Sir)Document9 pagesNon-Linear Pharmacokinetics: Biopharmaceutics (Ashik Sir)Hasan Al MasudNo ratings yet
- GSCL Diferences Kinetic EnzymeDocument3 pagesGSCL Diferences Kinetic Enzymejoenni hansNo ratings yet
- 9.07 Theories of Reaction Rates PDFDocument6 pages9.07 Theories of Reaction Rates PDFAbdelfattah Mohamed OufNo ratings yet
- Biochem ConceptsDocument9 pagesBiochem Conceptsdanena88No ratings yet
- Exam II ReviewDocument9 pagesExam II ReviewShira ChoiNo ratings yet
- Mechaelis-Menten KineticsDocument7 pagesMechaelis-Menten KineticsfayeNo ratings yet
- The Molecular Symmetry GroupDocument24 pagesThe Molecular Symmetry GroupWippetsxzNo ratings yet
- Chapter 6 Parallel DC CircuitsDocument72 pagesChapter 6 Parallel DC CircuitsAbdullrahman Al-ShammaaNo ratings yet
- Mikales Menten Great With Green Book AlsoDocument8 pagesMikales Menten Great With Green Book AlsoMOSES MILLERNo ratings yet
- Facts and PQ Unit-IIIDocument9 pagesFacts and PQ Unit-IIIAniket ANo ratings yet
- A Steverson Chapter 11 Pages 416-468Document54 pagesA Steverson Chapter 11 Pages 416-468Gerson NetoNo ratings yet
- Fundamentals of Electrical Circuits AnalysisDocument8 pagesFundamentals of Electrical Circuits AnalysisArcturus VoidNo ratings yet
- Effect of Battery Energy Storage System On Load Frequency Control Considering Governor Deadband and Generation Rate ConstraintDocument7 pagesEffect of Battery Energy Storage System On Load Frequency Control Considering Governor Deadband and Generation Rate ConstraintzahidNo ratings yet
- Ch06 QuizDocument5 pagesCh06 QuizcatlovesfoodNo ratings yet
- LAB 3 - NewDocument5 pagesLAB 3 - Newzaaa asdadsasdaNo ratings yet
- Kuzmic KobsTight 20220825 001Document7 pagesKuzmic KobsTight 20220825 001Petr KuzmicNo ratings yet
- Enzyme Kinetics: CHEM 3501 The University of Memphis Foundations of Bioorganic Chemistry Lab Department of ChemistryDocument6 pagesEnzyme Kinetics: CHEM 3501 The University of Memphis Foundations of Bioorganic Chemistry Lab Department of ChemistryMohammad AliNo ratings yet
- Midnight HWDocument3 pagesMidnight HWDont MeNo ratings yet
- Atkins EnzimasDocument5 pagesAtkins EnzimasConstanza Espinoza LaraNo ratings yet
- Derivations OF: Enzyme Kinetics (I)Document20 pagesDerivations OF: Enzyme Kinetics (I)Sabari Krishnan B B100% (1)
- Enzmology RevisionDocument8 pagesEnzmology RevisionRyan Fortune AludaNo ratings yet
- Lab 10 PDFDocument3 pagesLab 10 PDFFaiyaz ShaikhNo ratings yet
- Hypervariable Regions: Nature of Antigen-Antibody Reactions Lock and Key ConceptDocument7 pagesHypervariable Regions: Nature of Antigen-Antibody Reactions Lock and Key ConceptVenkata Ramana Murthy VasupilliNo ratings yet
- Capacitor Placement in Unbalanced Distribution Networks To Minimize Reactive and Unbalance Power LossesDocument8 pagesCapacitor Placement in Unbalanced Distribution Networks To Minimize Reactive and Unbalance Power LossesJones LennonNo ratings yet
- Chapter11-Two Ways AnovaDocument26 pagesChapter11-Two Ways AnovaRaluca NegruNo ratings yet
- Reviews in Computational ChemistryFrom EverandReviews in Computational ChemistryAbby L. ParrillNo ratings yet
- On The Integration of in Silico Drug Design Methods For Drug RepurposingDocument7 pagesOn The Integration of in Silico Drug Design Methods For Drug RepurposingdarmaNo ratings yet
- Indian Company ListDocument256 pagesIndian Company ListRaj SaNo ratings yet
- Mencari Selisih (1) EDITDocument426 pagesMencari Selisih (1) EDITlukiharjantiNo ratings yet
- Journal June 2019Document36 pagesJournal June 2019rammvr05No ratings yet
- Nursing Pharmacology BSN2Document9 pagesNursing Pharmacology BSN2Ria Angely PanagaNo ratings yet
- Presentation Adverse Drug ReactionDocument16 pagesPresentation Adverse Drug ReactionAG STXLEZNo ratings yet
- Anda - of AndaDocument46 pagesAnda - of AndaTarikNo ratings yet
- Question Paper DSSSB Phamacist April 2015Document29 pagesQuestion Paper DSSSB Phamacist April 2015Sahil KumarNo ratings yet
- BCS ClassDocument8 pagesBCS Classrina wijayantiNo ratings yet
- Handout On Prescription WritingDocument10 pagesHandout On Prescription WritingAbdiNo ratings yet
- Biopharmaceutics Supplemental NotesDocument19 pagesBiopharmaceutics Supplemental NotesChristine Joy De Ocampo0% (1)
- Inactive Ingredient Search For Approved Dhtghrug ProductsDocument2 pagesInactive Ingredient Search For Approved Dhtghrug ProductsDang Chi CongNo ratings yet
- Daftar Pustaka SterilDocument3 pagesDaftar Pustaka SterilNurhayati Nasrid BobaNo ratings yet
- Marketing ExamDocument42 pagesMarketing ExamMarianne CruzNo ratings yet
- HNMC CV Formulary PDFDocument129 pagesHNMC CV Formulary PDFAnonymous F1xtU2RNo ratings yet
- CTD Dossier Preparation: K. Srikantha Reddy K. Srikantha ReddyDocument46 pagesCTD Dossier Preparation: K. Srikantha Reddy K. Srikantha Reddyfadli100% (1)
- Nfi 2011Document804 pagesNfi 2011indmale_0070% (1)
- Anticancer DrugsDocument6 pagesAnticancer DrugsmidhunNo ratings yet
- Receptor Profile of Selected Anti-Psychotic Drugs: Lovely Liana Clapis Sittie Rahani CanacanDocument42 pagesReceptor Profile of Selected Anti-Psychotic Drugs: Lovely Liana Clapis Sittie Rahani CanacanKevin ChanNo ratings yet
- 2 TIVA - TCI RevisionDocument25 pages2 TIVA - TCI RevisionMora GalliNo ratings yet
- Drug Development and Regulatory Abbreviation SheetDocument16 pagesDrug Development and Regulatory Abbreviation SheetAdamNo ratings yet
- Hetero Corp Presentation 2016Document23 pagesHetero Corp Presentation 2016Evan TanswariNo ratings yet
- Cefuroxime 1st PDF TRIAL PDFDocument1 pageCefuroxime 1st PDF TRIAL PDFRyanNo ratings yet
- Daftar Inventaris Buku FFDocument71 pagesDaftar Inventaris Buku FFcakra gilingNo ratings yet
- Decreased Warfarin Effect After Initiation of High-Protein, Low-Carbohydrate DietsDocument5 pagesDecreased Warfarin Effect After Initiation of High-Protein, Low-Carbohydrate DietsDinaNo ratings yet
- Laporan DakDocument168 pagesLaporan DaknurhasanahNo ratings yet
- ACTD Administrative DataDocument14 pagesACTD Administrative DataWilliam ChandraNo ratings yet
- Orally Disintegrating Tablets: A Novel Approach For MedicationDocument8 pagesOrally Disintegrating Tablets: A Novel Approach For MedicationyNo ratings yet
- Pharmaceutical Care vs. Clinical PharmacyDocument2 pagesPharmaceutical Care vs. Clinical PharmacyKaren Park100% (1)