Download as docx, pdf, or txt
You might also like
- Maturski Rad Hrskavicave Ribe Raze I Ajkule FINALNODocument56 pagesMaturski Rad Hrskavicave Ribe Raze I Ajkule FINALNOBerinaVrhovac100% (1)
- Poreecaji Acidobazne Ravnoteze 2008Document73 pagesPoreecaji Acidobazne Ravnoteze 2008Natalija StamenkovicNo ratings yet
- Bioloski Znacaj PHDocument12 pagesBioloski Znacaj PHBojan KojicicNo ratings yet
- Poremećaji Acido Bazne Ravnoteže 1Document64 pagesPoremećaji Acido Bazne Ravnoteže 1Marija ZivkovicNo ratings yet
- D KEMIJSKA SVOJSTVA ORGANSKIH SPOJEVA S KISIKOMDocument14 pagesD KEMIJSKA SVOJSTVA ORGANSKIH SPOJEVA S KISIKOMlucius maharalNo ratings yet
- Gradja VirusaDocument13 pagesGradja VirusaGordanaStevovicMladenovicNo ratings yet
- Ekologija Zivotinja 40 OKDocument84 pagesEkologija Zivotinja 40 OKAlex AnnieNo ratings yet
- IV Raz 8 KARBOKSILNE KISELINE 2020 +++Document6 pagesIV Raz 8 KARBOKSILNE KISELINE 2020 +++Egov AlteroNo ratings yet
- KožaDocument4 pagesKožaasko91444No ratings yet
- Multipli AleliDocument29 pagesMultipli AleliЖИКА ПАУНКОВИЋNo ratings yet
- Biohemija - Metabolizam I Ugljeni HidratiDocument17 pagesBiohemija - Metabolizam I Ugljeni HidratiAleksandarNo ratings yet
- Tablica IonaDocument2 pagesTablica IonaMarkoNo ratings yet
- HromatografijaDocument11 pagesHromatografijadubravkadjNo ratings yet
- KiselineDocument1 pageKiselineRafaela ČavlinaNo ratings yet
- Etologija PitanjaDocument5 pagesEtologija PitanjaAdnaNo ratings yet
- Poglavlje1-Pigmenti Za DisanjesdDocument80 pagesPoglavlje1-Pigmenti Za DisanjesdEmina Isic ZovkicNo ratings yet
- Karboksilne KiselineDocument7 pagesKarboksilne KiselineAndrea BabogredacNo ratings yet
- Karboksilne KiselineDocument68 pagesKarboksilne KiselineLaura100% (1)
- Praktikum Iz Hemije Za Studente Medicine - Medicinska Biohemija I HemijaDocument64 pagesPraktikum Iz Hemije Za Studente Medicine - Medicinska Biohemija I HemijaBožidar ĆaćićNo ratings yet
- Bilješka o PiscuDocument5 pagesBilješka o PiscuAlisaNo ratings yet
- H.mujovic - Prekid TrudnoceDocument30 pagesH.mujovic - Prekid TrudnoceTeodor MilanovićNo ratings yet
- Higijena RukuDocument15 pagesHigijena RukuAmer NurkovicNo ratings yet
- Drugi Razred Neorganska Hemija SkripteDocument54 pagesDrugi Razred Neorganska Hemija SkripteKirilKocevskiNo ratings yet
- Termalno Zagadjenje Vode 2003Document13 pagesTermalno Zagadjenje Vode 2003Milena DalmacijaNo ratings yet
- 13-Karboksilne KiselineDocument63 pages13-Karboksilne KiselineNihad HasanovićNo ratings yet
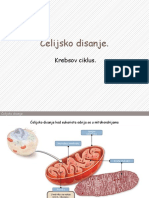
- Krebsov Ciklus. Elektron - Transportni LanacDocument18 pagesKrebsov Ciklus. Elektron - Transportni LanacGoranNo ratings yet
- Uvod U Vet, SeminarskiDocument19 pagesUvod U Vet, SeminarskiMarkoP2861No ratings yet
- Ekologija 2016 17Document8 pagesEkologija 2016 17Kljucanin SenaidNo ratings yet
- Određivanje Fizičkih I Hemijskih Osobina VodeDocument8 pagesOdređivanje Fizičkih I Hemijskih Osobina VodeNikola Belobaba50% (2)
- Voda Kao RastvaracDocument4 pagesVoda Kao RastvaracKarijaz Vseslav100% (1)
- LipidiDocument38 pagesLipidiDači KalenderNo ratings yet
- 2276 Sociologija Moral I Obicaji SRB 15strDocument16 pages2276 Sociologija Moral I Obicaji SRB 15straska1990No ratings yet
- Ugljeni HidratiDocument11 pagesUgljeni HidratiNikola SvorcanNo ratings yet
- Kapilarne PojaveDocument6 pagesKapilarne PojaveMunevera OriflameNo ratings yet
- ElektrolitiDocument14 pagesElektrolitiNikola BarbulovićNo ratings yet
- Biologija IIDocument2 pagesBiologija IIastufisNo ratings yet
- 2 Cas TRANSPORT KROZ CELIJSKU MEMBRANU PASIVAN TRANSPORT-25719130702901117558Document6 pages2 Cas TRANSPORT KROZ CELIJSKU MEMBRANU PASIVAN TRANSPORT-25719130702901117558Ilija100% (1)
- Uvod U Neorgansku HemijuDocument1 pageUvod U Neorgansku HemijuBiljana PenjinNo ratings yet
- Zloupotreba Psihoaktivnih SupstanciDocument22 pagesZloupotreba Psihoaktivnih SupstanciMiriam FedericoNo ratings yet
- Glasovne PromeneDocument46 pagesGlasovne PromeneАна ЛазовићNo ratings yet
- Kardiovaskularni SistemDocument36 pagesKardiovaskularni SistemGoran StamenkovicNo ratings yet
- Morfologija TestDocument7 pagesMorfologija TestŠišmišova DevojkaNo ratings yet
- Karboksilne Kiseline I Derivati Karboksilnih KiselinaDocument24 pagesKarboksilne Kiseline I Derivati Karboksilnih KiselinaPravnikNPNo ratings yet
- Biologija - Genetika 1Document4 pagesBiologija - Genetika 1Aleksandar MišićNo ratings yet
- Kontrolni Zadatak - A Grupa DS PrvaciDocument2 pagesKontrolni Zadatak - A Grupa DS PrvaciMiroslava100% (1)
- СТАНОВНИШТВО И ДРУШТВОDocument4 pagesСТАНОВНИШТВО И ДРУШТВОJovanaNo ratings yet
- Direktno Merenje Po MasiDocument5 pagesDirektno Merenje Po MasiVesna Milanović SimičićNo ratings yet
- Test Pitanja Bolesti Malih ZivotinjaDocument4 pagesTest Pitanja Bolesti Malih ZivotinjaStanislava KanjevacNo ratings yet
- Угрожене врсте животињаDocument22 pagesУгрожене врсте животињаJelena Ognjanovic0% (1)
- LipidiDocument66 pagesLipidiTijana S. PetkovićNo ratings yet
- M-3 Lekarski Pregled 1314-2Document1 pageM-3 Lekarski Pregled 1314-2MSc Kostic MilosNo ratings yet
- Indirektna JodimetrijaDocument3 pagesIndirektna Jodimetrijaalma456100% (1)
- Aminokiseline I Peptidna VezaDocument11 pagesAminokiseline I Peptidna VezaANo ratings yet
- Estetski TurizamDocument11 pagesEstetski TurizamBojan AnticNo ratings yet
- Vektori U Koordinatnom SustavuDocument7 pagesVektori U Koordinatnom SustavuzdravkoNo ratings yet
- Hemija Drveta 2. TestDocument32 pagesHemija Drveta 2. TestMemi100% (1)
- 3158 Vodic Za Doktore Veterinarske Medicine PDFDocument48 pages3158 Vodic Za Doktore Veterinarske Medicine PDFSlobodan MiloševićNo ratings yet
- AlkoholiDocument18 pagesAlkoholiAneri88No ratings yet
- Acido BaznaDocument4 pagesAcido BaznaabarksasNo ratings yet
- BiohemijaDocument7 pagesBiohemijaRobert VinsonNo ratings yet