
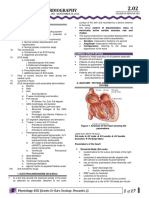
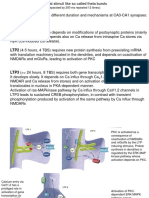
Synaptic Depression: Many Different Possible Mechanisms
Synaptic Depression: Many Different Possible Mechanisms
You might also like
- 2013 Spyder ST Owners ManualDocument171 pages2013 Spyder ST Owners ManualMelvin Quezada67% (6)
- Bipn 105 Final Study GuideDocument7 pagesBipn 105 Final Study Guideapi-307388626No ratings yet
- Hvac PM Service Checklist ContractorDocument1 pageHvac PM Service Checklist Contractorudupiganesh3069100% (2)
- Talal Asad - Anthropology and The Colonial EncounterDocument4 pagesTalal Asad - Anthropology and The Colonial EncounterTania Saha100% (1)
- HubSpot: Inbound Marketing and Web 2.0Document3 pagesHubSpot: Inbound Marketing and Web 2.0test100% (1)
- Detailed Lesson Plan in English IV - COMPOUND SENTENCEDocument12 pagesDetailed Lesson Plan in English IV - COMPOUND SENTENCEMadhie Gre Dela Provi94% (16)
- PDF Document 22Document1 pagePDF Document 22guzelinsan727No ratings yet
- Debanne Et Al-1998-The Journal of PhysiologyDocument12 pagesDebanne Et Al-1998-The Journal of PhysiologyGabriel HerreraNo ratings yet
- Repetitve Nerve Stimulation (RNS) : By: Syed Irshad Murtaza Neurophysiology Dept AKUH Karachi Date:12-06-2013Document33 pagesRepetitve Nerve Stimulation (RNS) : By: Syed Irshad Murtaza Neurophysiology Dept AKUH Karachi Date:12-06-2013Raghu NadhNo ratings yet
- Synaptic IntegrationDocument5 pagesSynaptic IntegrationryanNo ratings yet
- KORN Et Al - Spontaneous Quantal Currents in A Central Neuron Match Predictions From Binominal Analysis of Evoked ResponsesDocument5 pagesKORN Et Al - Spontaneous Quantal Currents in A Central Neuron Match Predictions From Binominal Analysis of Evoked Responsesfernando_mart859243No ratings yet
- Lecture 8ADocument17 pagesLecture 8AAbdullah EwaisNo ratings yet
- Section 2: Signal Transmission Between The NeuronsDocument123 pagesSection 2: Signal Transmission Between The NeuronsAna Crigan VlasNo ratings yet
- Jphysiol00958 0128Document26 pagesJphysiol00958 0128marielaalejandrarNo ratings yet
- Synapses: Department of Mathematical SciencesDocument7 pagesSynapses: Department of Mathematical SciencesilincaNo ratings yet
- Zheng 2006 NatNeuro Bursting Intrinsic Retina PDFDocument9 pagesZheng 2006 NatNeuro Bursting Intrinsic Retina PDFSimón Salgado MuñozNo ratings yet
- Buzsaki02 Nikolaus ExzerptDocument7 pagesBuzsaki02 Nikolaus ExzerptsiamakNo ratings yet
- Membrane Stretch Affects Gating Modes of A Skeletal Muscle Sodium ChannelDocument17 pagesMembrane Stretch Affects Gating Modes of A Skeletal Muscle Sodium ChannelMichael MaierNo ratings yet
- Effects of Low Concentrations of 4-Aminopyridine On CA1 Pyramidal Cells of The HippocampusDocument18 pagesEffects of Low Concentrations of 4-Aminopyridine On CA1 Pyramidal Cells of The HippocampusJortegloriaNo ratings yet
- Paf in LTPDocument6 pagesPaf in LTPKapil SoniNo ratings yet
- Summation (Neurophysiology) : Wiki Loves Monuments: Photograph A Monument, Help Wikipedia and Win!Document9 pagesSummation (Neurophysiology) : Wiki Loves Monuments: Photograph A Monument, Help Wikipedia and Win!Jennie KimNo ratings yet
- Michael R. Guevara and Habo J. Jongsma - Phase Resetting in A Model of Sinoatrial Nodal Membrane: Ionic and Topological AspectsDocument14 pagesMichael R. Guevara and Habo J. Jongsma - Phase Resetting in A Model of Sinoatrial Nodal Membrane: Ionic and Topological AspectsLuammmNo ratings yet
- L2 Neurons and Neuronal NetworksDocument4 pagesL2 Neurons and Neuronal NetworksmoshlingmomoNo ratings yet
- SynapseDocument48 pagesSynapseSaurabh KrishnaNo ratings yet
- Synaptic Transmission 4Document36 pagesSynaptic Transmission 4stehephNo ratings yet
- Marco Fuenzalida Et Al - Changes of The EPSP Waveform Regulate The Temporal Window For Spike-Timing-Dependent PlasticityDocument9 pagesMarco Fuenzalida Et Al - Changes of The EPSP Waveform Regulate The Temporal Window For Spike-Timing-Dependent PlasticityFedrmNo ratings yet
- Cellular Neuroscience (207) : Ian ParkerDocument12 pagesCellular Neuroscience (207) : Ian ParkerAndreyIvanovNo ratings yet
- tmpABFB TMPDocument11 pagestmpABFB TMPFrontiersNo ratings yet
- Takuya Yasui Et Al - Dynamic Synapses As Archives of Synaptic History: State-Dependent Redistribution of Synaptic Efficacy in The Rat Hippocampal CA1Document18 pagesTakuya Yasui Et Al - Dynamic Synapses As Archives of Synaptic History: State-Dependent Redistribution of Synaptic Efficacy in The Rat Hippocampal CA1FedrmNo ratings yet
- CH5 THE SYNAPSE AND ACTION POTENTIAL GENERATION ON THE POSTSYNAPTIC MEMRANE NewDocument4 pagesCH5 THE SYNAPSE AND ACTION POTENTIAL GENERATION ON THE POSTSYNAPTIC MEMRANE NewoguzNo ratings yet
- 05 02 Rabbit Nerve ImpulseDocument10 pages05 02 Rabbit Nerve ImpulseTan Hing LeeNo ratings yet
- ModelSystem Synapse NeurotransmittersDocument41 pagesModelSystem Synapse NeurotransmittersTapan Kumar PalNo ratings yet
- NROSCI 1012 - Lecture 22Document4 pagesNROSCI 1012 - Lecture 22HonzaNo ratings yet
- TMP 9 DA2Document11 pagesTMP 9 DA2FrontiersNo ratings yet
- Carbamazepine-Induced Suppression of Repetitive Firing in CA1 Pyramidal Neurons Is Greater in The Dorsal Hippocampus Than The Ventral HippocampusDocument10 pagesCarbamazepine-Induced Suppression of Repetitive Firing in CA1 Pyramidal Neurons Is Greater in The Dorsal Hippocampus Than The Ventral HippocampusGading AuroraNo ratings yet
- Lin Etal 2003Document9 pagesLin Etal 2003林瑜璿No ratings yet
- Synapses and Synaptic Transmission: Dr. Eman El Eter Physiology Dep. College of Medicine KSUDocument43 pagesSynapses and Synaptic Transmission: Dr. Eman El Eter Physiology Dep. College of Medicine KSUraanja2No ratings yet
- Cold Spring Harb Perspect Biol-2012-Blakely-A005595Document25 pagesCold Spring Harb Perspect Biol-2012-Blakely-A005595Md. Ahsan-Ul BariNo ratings yet
- NROSCI 1012 - Lecture 24Document4 pagesNROSCI 1012 - Lecture 24HonzaNo ratings yet
- Brain Res 1977 AghajanianDocument8 pagesBrain Res 1977 Aghajanianndyfk54dpgNo ratings yet
- Novel Strategy For Measuring Creatine Kinase Reaction Rate in The in Vivo HeartDocument10 pagesNovel Strategy For Measuring Creatine Kinase Reaction Rate in The in Vivo HeartYoung-Hoon SungNo ratings yet
- Synaptic TransmissionDocument17 pagesSynaptic TransmissionbnmjgcNo ratings yet
- Action Potential Generation in Cardiac MuscleDocument10 pagesAction Potential Generation in Cardiac MuscleAhmad HaNo ratings yet
- Biophysj00107 0239Document6 pagesBiophysj00107 0239aydinmemmedov9009No ratings yet
- Cardiac Electrophysiology Basics (RL-204-Tn)Document5 pagesCardiac Electrophysiology Basics (RL-204-Tn)Navojit ChowdhuryNo ratings yet
- ECG of The Month: DiscussionDocument2 pagesECG of The Month: DiscussionKrlÖz Ändrz Hërëdîa ÖpzNo ratings yet
- Christian Ducreux and Jean-Jacques Puizillout - A-Current Modifies The Spike of C-Type Neurones in The Rabbit Nodose GanglionDocument13 pagesChristian Ducreux and Jean-Jacques Puizillout - A-Current Modifies The Spike of C-Type Neurones in The Rabbit Nodose GanglionFedrmNo ratings yet
- Cardiac MuscleDocument62 pagesCardiac Musclegxw5fw99zsNo ratings yet
- Cordycepin - CA1 Membrane HyperpolarizationDocument5 pagesCordycepin - CA1 Membrane Hyperpolarizationttqnhu.rhmNo ratings yet
- Calcium Control of Neurotransmitter Release PDFDocument15 pagesCalcium Control of Neurotransmitter Release PDFNisa Yulianti SuprahmanNo ratings yet
- Calcium Control of Neurotransmitter ReleaseDocument15 pagesCalcium Control of Neurotransmitter ReleaseNisa Yulianti SuprahmanNo ratings yet
- Dopaminergic Modulation of Short-Term Synaptic Plasticity at Striatal Inhibitory SynapsesDocument6 pagesDopaminergic Modulation of Short-Term Synaptic Plasticity at Striatal Inhibitory SynapsesMaría Celeste Sánchez SugíaNo ratings yet
- L1 Neurons and Neuronal NetworksDocument4 pagesL1 Neurons and Neuronal NetworksmoshlingmomoNo ratings yet
- Cunha-Reis BrJPharm143 (06) 733 (2004)Document12 pagesCunha-Reis BrJPharm143 (06) 733 (2004)dcrrufaNo ratings yet
- Synaptic Transmission: Transmission at The Synapses May BeDocument13 pagesSynaptic Transmission: Transmission at The Synapses May Beramadan100% (3)
- Cardiovascular Physiology Review QuestionsDocument44 pagesCardiovascular Physiology Review QuestionsTim100% (1)
- 2 02-Electrocardiography PDFDocument17 pages2 02-Electrocardiography PDFMiguel DomingoNo ratings yet
- Lin 1993Document13 pagesLin 1993Stress LessNo ratings yet
- QuizletDocument7 pagesQuizletSarath kumarNo ratings yet
- BIPN 100, Mammalian Physiology I - Solutions To Problem Set #1Document5 pagesBIPN 100, Mammalian Physiology I - Solutions To Problem Set #1whateverpplsNo ratings yet
- RAT IN: Bicuculline Potential (EPSP) - Relationship EPSP Amplitude AmplitudeDocument14 pagesRAT IN: Bicuculline Potential (EPSP) - Relationship EPSP Amplitude AmplitudeFedrmNo ratings yet
- CardiacDocument32 pagesCardiacAlli OmerNo ratings yet
- A Most Important Question: Click Here To ViewDocument1 pageA Most Important Question: Click Here To ViewLwin Maung Maung ThikeNo ratings yet
- Neurobiology 4 - A - Synaptic PlasticityDocument26 pagesNeurobiology 4 - A - Synaptic PlasticitymolinakevinalejandroNo ratings yet
- Computational NeuroendocrinologyFrom EverandComputational NeuroendocrinologyDuncan J. MacGregorNo ratings yet
- Neurobiology Assignment PDFDocument2 pagesNeurobiology Assignment PDFThe Panda EntertainerNo ratings yet
- 24 Short - Term - Depression - TC FFI - Optional PDFDocument9 pages24 Short - Term - Depression - TC FFI - Optional PDFThe Panda EntertainerNo ratings yet
- Stimulation of Neuronal Activity With LightDocument29 pagesStimulation of Neuronal Activity With LightThe Panda EntertainerNo ratings yet
- 25 Learning - Memory1 PDFDocument28 pages25 Learning - Memory1 PDFThe Panda EntertainerNo ratings yet
- LTP1 ( 2 Hours, 1 TBS) Depends On Modifications of Postsynaptic Proteins (MainlyDocument12 pagesLTP1 ( 2 Hours, 1 TBS) Depends On Modifications of Postsynaptic Proteins (MainlyThe Panda EntertainerNo ratings yet
- Mechanisms of Short-Term Synaptic PlasticityDocument13 pagesMechanisms of Short-Term Synaptic PlasticityThe Panda EntertainerNo ratings yet
- 21 Synaptic Plasticity - Short - Term PDFDocument19 pages21 Synaptic Plasticity - Short - Term PDFThe Panda EntertainerNo ratings yet
- 28 Optical - Techniques PDFDocument28 pages28 Optical - Techniques PDFThe Panda EntertainerNo ratings yet
- 20 Synaptic Transmission - Vel - Ca - Nano Vs Microdomains PDFDocument23 pages20 Synaptic Transmission - Vel - Ca - Nano Vs Microdomains PDFThe Panda EntertainerNo ratings yet
- Name:: Nyoman Gede Abhyasa Class: X Mipa 2 Absent: 25 AnswersDocument2 pagesName:: Nyoman Gede Abhyasa Class: X Mipa 2 Absent: 25 AnswersRetrify - Random ContentNo ratings yet
- Dsm-5 Icd-10 HandoutDocument107 pagesDsm-5 Icd-10 HandoutAakanksha Verma100% (1)
- Themelis Ulloa LandfillDocument15 pagesThemelis Ulloa LandfillHenry Bagus WicaksonoNo ratings yet
- Sip 2019-2022Document4 pagesSip 2019-2022jein_am97% (29)
- September 23, Infer The Purpose of The Poem Listened ToDocument4 pagesSeptember 23, Infer The Purpose of The Poem Listened ToLouelle GonzalesNo ratings yet
- Lawrance Africa Imagined in The Spanish Renaissance - Henry Thomas LectureDocument20 pagesLawrance Africa Imagined in The Spanish Renaissance - Henry Thomas LecturejlawranceNo ratings yet
- Prolog Programs Are A Collection of FactsDocument23 pagesProlog Programs Are A Collection of FactsAbni booNo ratings yet
- ResearchDocument38 pagesResearchHelen McClintockNo ratings yet
- Catalog de Produse - BTRDocument62 pagesCatalog de Produse - BTRdaliproiectareNo ratings yet
- Act 1 Almeyda JTLDocument2 pagesAct 1 Almeyda JTLAltairNo ratings yet
- Marik ServitorsDocument20 pagesMarik ServitorsDWNloader100% (2)
- Taylor Swift LyricsDocument2 pagesTaylor Swift LyricsElsie DomeNo ratings yet
- APA-AP - Activity 3-Emerging Ethical and Policy Issues of STSDocument36 pagesAPA-AP - Activity 3-Emerging Ethical and Policy Issues of STSJOHN MARTIN APA-APNo ratings yet
- ContinueDocument3 pagesContinueJúÑi ØrNo ratings yet
- Arlon 38NDocument4 pagesArlon 38NRavindrakumar ParvathiniNo ratings yet
- 501-453 Universal Cable GalndsDocument1 page501-453 Universal Cable Galndsmeribout adelNo ratings yet
- Group 24 ReportDocument48 pagesGroup 24 ReportVishwanath GroupsNo ratings yet
- Glass Configurator Datasheet 2023 03 27Document1 pageGlass Configurator Datasheet 2023 03 27Satrio PrakosoNo ratings yet
- Ethical Considerations Dissertation ExampleDocument7 pagesEthical Considerations Dissertation ExampleBuyPapersOnlineCanada100% (1)
- TakeawayDocument6 pagesTakeawayWilman VasquezNo ratings yet
- Chapter 1 3 THESISDocument64 pagesChapter 1 3 THESISLyka Cartagena-Pandili Manalo-CabelloNo ratings yet
- Drama Bahasa InggrisDocument24 pagesDrama Bahasa InggrisCahyo Yang bahagiaNo ratings yet
- Indias Gold Market Reform and GrowthDocument149 pagesIndias Gold Market Reform and GrowthtmeygmvzjfnkqcwhgpNo ratings yet
- DAPA Teams Spider DiagramDocument2 pagesDAPA Teams Spider DiagramEduardoJaimeNo ratings yet
- Omni-Directional Mobile Robot With Mecanum WheelDocument6 pagesOmni-Directional Mobile Robot With Mecanum WheelTRUNG QUOC LENo ratings yet

































































































Download as pdf or txt
You might also like
- 2013 Spyder ST Owners ManualDocument171 pages2013 Spyder ST Owners ManualMelvin Quezada67% (6)
- Bipn 105 Final Study GuideDocument7 pagesBipn 105 Final Study Guideapi-307388626No ratings yet
- Hvac PM Service Checklist ContractorDocument1 pageHvac PM Service Checklist Contractorudupiganesh3069100% (2)
- Talal Asad - Anthropology and The Colonial EncounterDocument4 pagesTalal Asad - Anthropology and The Colonial EncounterTania Saha100% (1)
- HubSpot: Inbound Marketing and Web 2.0Document3 pagesHubSpot: Inbound Marketing and Web 2.0test100% (1)
- Detailed Lesson Plan in English IV - COMPOUND SENTENCEDocument12 pagesDetailed Lesson Plan in English IV - COMPOUND SENTENCEMadhie Gre Dela Provi94% (16)
- PDF Document 22Document1 pagePDF Document 22guzelinsan727No ratings yet
- Debanne Et Al-1998-The Journal of PhysiologyDocument12 pagesDebanne Et Al-1998-The Journal of PhysiologyGabriel HerreraNo ratings yet
- Repetitve Nerve Stimulation (RNS) : By: Syed Irshad Murtaza Neurophysiology Dept AKUH Karachi Date:12-06-2013Document33 pagesRepetitve Nerve Stimulation (RNS) : By: Syed Irshad Murtaza Neurophysiology Dept AKUH Karachi Date:12-06-2013Raghu NadhNo ratings yet
- Synaptic IntegrationDocument5 pagesSynaptic IntegrationryanNo ratings yet
- KORN Et Al - Spontaneous Quantal Currents in A Central Neuron Match Predictions From Binominal Analysis of Evoked ResponsesDocument5 pagesKORN Et Al - Spontaneous Quantal Currents in A Central Neuron Match Predictions From Binominal Analysis of Evoked Responsesfernando_mart859243No ratings yet
- Lecture 8ADocument17 pagesLecture 8AAbdullah EwaisNo ratings yet
- Section 2: Signal Transmission Between The NeuronsDocument123 pagesSection 2: Signal Transmission Between The NeuronsAna Crigan VlasNo ratings yet
- Jphysiol00958 0128Document26 pagesJphysiol00958 0128marielaalejandrarNo ratings yet
- Synapses: Department of Mathematical SciencesDocument7 pagesSynapses: Department of Mathematical SciencesilincaNo ratings yet
- Zheng 2006 NatNeuro Bursting Intrinsic Retina PDFDocument9 pagesZheng 2006 NatNeuro Bursting Intrinsic Retina PDFSimón Salgado MuñozNo ratings yet
- Buzsaki02 Nikolaus ExzerptDocument7 pagesBuzsaki02 Nikolaus ExzerptsiamakNo ratings yet
- Membrane Stretch Affects Gating Modes of A Skeletal Muscle Sodium ChannelDocument17 pagesMembrane Stretch Affects Gating Modes of A Skeletal Muscle Sodium ChannelMichael MaierNo ratings yet
- Effects of Low Concentrations of 4-Aminopyridine On CA1 Pyramidal Cells of The HippocampusDocument18 pagesEffects of Low Concentrations of 4-Aminopyridine On CA1 Pyramidal Cells of The HippocampusJortegloriaNo ratings yet
- Paf in LTPDocument6 pagesPaf in LTPKapil SoniNo ratings yet
- Summation (Neurophysiology) : Wiki Loves Monuments: Photograph A Monument, Help Wikipedia and Win!Document9 pagesSummation (Neurophysiology) : Wiki Loves Monuments: Photograph A Monument, Help Wikipedia and Win!Jennie KimNo ratings yet
- Michael R. Guevara and Habo J. Jongsma - Phase Resetting in A Model of Sinoatrial Nodal Membrane: Ionic and Topological AspectsDocument14 pagesMichael R. Guevara and Habo J. Jongsma - Phase Resetting in A Model of Sinoatrial Nodal Membrane: Ionic and Topological AspectsLuammmNo ratings yet
- L2 Neurons and Neuronal NetworksDocument4 pagesL2 Neurons and Neuronal NetworksmoshlingmomoNo ratings yet
- SynapseDocument48 pagesSynapseSaurabh KrishnaNo ratings yet
- Synaptic Transmission 4Document36 pagesSynaptic Transmission 4stehephNo ratings yet
- Marco Fuenzalida Et Al - Changes of The EPSP Waveform Regulate The Temporal Window For Spike-Timing-Dependent PlasticityDocument9 pagesMarco Fuenzalida Et Al - Changes of The EPSP Waveform Regulate The Temporal Window For Spike-Timing-Dependent PlasticityFedrmNo ratings yet
- Cellular Neuroscience (207) : Ian ParkerDocument12 pagesCellular Neuroscience (207) : Ian ParkerAndreyIvanovNo ratings yet
- tmpABFB TMPDocument11 pagestmpABFB TMPFrontiersNo ratings yet
- Takuya Yasui Et Al - Dynamic Synapses As Archives of Synaptic History: State-Dependent Redistribution of Synaptic Efficacy in The Rat Hippocampal CA1Document18 pagesTakuya Yasui Et Al - Dynamic Synapses As Archives of Synaptic History: State-Dependent Redistribution of Synaptic Efficacy in The Rat Hippocampal CA1FedrmNo ratings yet
- CH5 THE SYNAPSE AND ACTION POTENTIAL GENERATION ON THE POSTSYNAPTIC MEMRANE NewDocument4 pagesCH5 THE SYNAPSE AND ACTION POTENTIAL GENERATION ON THE POSTSYNAPTIC MEMRANE NewoguzNo ratings yet
- 05 02 Rabbit Nerve ImpulseDocument10 pages05 02 Rabbit Nerve ImpulseTan Hing LeeNo ratings yet
- ModelSystem Synapse NeurotransmittersDocument41 pagesModelSystem Synapse NeurotransmittersTapan Kumar PalNo ratings yet
- NROSCI 1012 - Lecture 22Document4 pagesNROSCI 1012 - Lecture 22HonzaNo ratings yet
- TMP 9 DA2Document11 pagesTMP 9 DA2FrontiersNo ratings yet
- Carbamazepine-Induced Suppression of Repetitive Firing in CA1 Pyramidal Neurons Is Greater in The Dorsal Hippocampus Than The Ventral HippocampusDocument10 pagesCarbamazepine-Induced Suppression of Repetitive Firing in CA1 Pyramidal Neurons Is Greater in The Dorsal Hippocampus Than The Ventral HippocampusGading AuroraNo ratings yet
- Lin Etal 2003Document9 pagesLin Etal 2003林瑜璿No ratings yet
- Synapses and Synaptic Transmission: Dr. Eman El Eter Physiology Dep. College of Medicine KSUDocument43 pagesSynapses and Synaptic Transmission: Dr. Eman El Eter Physiology Dep. College of Medicine KSUraanja2No ratings yet
- Cold Spring Harb Perspect Biol-2012-Blakely-A005595Document25 pagesCold Spring Harb Perspect Biol-2012-Blakely-A005595Md. Ahsan-Ul BariNo ratings yet
- NROSCI 1012 - Lecture 24Document4 pagesNROSCI 1012 - Lecture 24HonzaNo ratings yet
- Brain Res 1977 AghajanianDocument8 pagesBrain Res 1977 Aghajanianndyfk54dpgNo ratings yet
- Novel Strategy For Measuring Creatine Kinase Reaction Rate in The in Vivo HeartDocument10 pagesNovel Strategy For Measuring Creatine Kinase Reaction Rate in The in Vivo HeartYoung-Hoon SungNo ratings yet
- Synaptic TransmissionDocument17 pagesSynaptic TransmissionbnmjgcNo ratings yet
- Action Potential Generation in Cardiac MuscleDocument10 pagesAction Potential Generation in Cardiac MuscleAhmad HaNo ratings yet
- Biophysj00107 0239Document6 pagesBiophysj00107 0239aydinmemmedov9009No ratings yet
- Cardiac Electrophysiology Basics (RL-204-Tn)Document5 pagesCardiac Electrophysiology Basics (RL-204-Tn)Navojit ChowdhuryNo ratings yet
- ECG of The Month: DiscussionDocument2 pagesECG of The Month: DiscussionKrlÖz Ändrz Hërëdîa ÖpzNo ratings yet
- Christian Ducreux and Jean-Jacques Puizillout - A-Current Modifies The Spike of C-Type Neurones in The Rabbit Nodose GanglionDocument13 pagesChristian Ducreux and Jean-Jacques Puizillout - A-Current Modifies The Spike of C-Type Neurones in The Rabbit Nodose GanglionFedrmNo ratings yet
- Cardiac MuscleDocument62 pagesCardiac Musclegxw5fw99zsNo ratings yet
- Cordycepin - CA1 Membrane HyperpolarizationDocument5 pagesCordycepin - CA1 Membrane Hyperpolarizationttqnhu.rhmNo ratings yet
- Calcium Control of Neurotransmitter Release PDFDocument15 pagesCalcium Control of Neurotransmitter Release PDFNisa Yulianti SuprahmanNo ratings yet
- Calcium Control of Neurotransmitter ReleaseDocument15 pagesCalcium Control of Neurotransmitter ReleaseNisa Yulianti SuprahmanNo ratings yet
- Dopaminergic Modulation of Short-Term Synaptic Plasticity at Striatal Inhibitory SynapsesDocument6 pagesDopaminergic Modulation of Short-Term Synaptic Plasticity at Striatal Inhibitory SynapsesMaría Celeste Sánchez SugíaNo ratings yet
- L1 Neurons and Neuronal NetworksDocument4 pagesL1 Neurons and Neuronal NetworksmoshlingmomoNo ratings yet
- Cunha-Reis BrJPharm143 (06) 733 (2004)Document12 pagesCunha-Reis BrJPharm143 (06) 733 (2004)dcrrufaNo ratings yet
- Synaptic Transmission: Transmission at The Synapses May BeDocument13 pagesSynaptic Transmission: Transmission at The Synapses May Beramadan100% (3)
- Cardiovascular Physiology Review QuestionsDocument44 pagesCardiovascular Physiology Review QuestionsTim100% (1)
- 2 02-Electrocardiography PDFDocument17 pages2 02-Electrocardiography PDFMiguel DomingoNo ratings yet
- Lin 1993Document13 pagesLin 1993Stress LessNo ratings yet
- QuizletDocument7 pagesQuizletSarath kumarNo ratings yet
- BIPN 100, Mammalian Physiology I - Solutions To Problem Set #1Document5 pagesBIPN 100, Mammalian Physiology I - Solutions To Problem Set #1whateverpplsNo ratings yet
- RAT IN: Bicuculline Potential (EPSP) - Relationship EPSP Amplitude AmplitudeDocument14 pagesRAT IN: Bicuculline Potential (EPSP) - Relationship EPSP Amplitude AmplitudeFedrmNo ratings yet
- CardiacDocument32 pagesCardiacAlli OmerNo ratings yet
- A Most Important Question: Click Here To ViewDocument1 pageA Most Important Question: Click Here To ViewLwin Maung Maung ThikeNo ratings yet
- Neurobiology 4 - A - Synaptic PlasticityDocument26 pagesNeurobiology 4 - A - Synaptic PlasticitymolinakevinalejandroNo ratings yet
- Computational NeuroendocrinologyFrom EverandComputational NeuroendocrinologyDuncan J. MacGregorNo ratings yet
- Neurobiology Assignment PDFDocument2 pagesNeurobiology Assignment PDFThe Panda EntertainerNo ratings yet
- 24 Short - Term - Depression - TC FFI - Optional PDFDocument9 pages24 Short - Term - Depression - TC FFI - Optional PDFThe Panda EntertainerNo ratings yet
- Stimulation of Neuronal Activity With LightDocument29 pagesStimulation of Neuronal Activity With LightThe Panda EntertainerNo ratings yet
- 25 Learning - Memory1 PDFDocument28 pages25 Learning - Memory1 PDFThe Panda EntertainerNo ratings yet
- LTP1 ( 2 Hours, 1 TBS) Depends On Modifications of Postsynaptic Proteins (MainlyDocument12 pagesLTP1 ( 2 Hours, 1 TBS) Depends On Modifications of Postsynaptic Proteins (MainlyThe Panda EntertainerNo ratings yet
- Mechanisms of Short-Term Synaptic PlasticityDocument13 pagesMechanisms of Short-Term Synaptic PlasticityThe Panda EntertainerNo ratings yet
- 21 Synaptic Plasticity - Short - Term PDFDocument19 pages21 Synaptic Plasticity - Short - Term PDFThe Panda EntertainerNo ratings yet
- 28 Optical - Techniques PDFDocument28 pages28 Optical - Techniques PDFThe Panda EntertainerNo ratings yet
- 20 Synaptic Transmission - Vel - Ca - Nano Vs Microdomains PDFDocument23 pages20 Synaptic Transmission - Vel - Ca - Nano Vs Microdomains PDFThe Panda EntertainerNo ratings yet
- Name:: Nyoman Gede Abhyasa Class: X Mipa 2 Absent: 25 AnswersDocument2 pagesName:: Nyoman Gede Abhyasa Class: X Mipa 2 Absent: 25 AnswersRetrify - Random ContentNo ratings yet
- Dsm-5 Icd-10 HandoutDocument107 pagesDsm-5 Icd-10 HandoutAakanksha Verma100% (1)
- Themelis Ulloa LandfillDocument15 pagesThemelis Ulloa LandfillHenry Bagus WicaksonoNo ratings yet
- Sip 2019-2022Document4 pagesSip 2019-2022jein_am97% (29)
- September 23, Infer The Purpose of The Poem Listened ToDocument4 pagesSeptember 23, Infer The Purpose of The Poem Listened ToLouelle GonzalesNo ratings yet
- Lawrance Africa Imagined in The Spanish Renaissance - Henry Thomas LectureDocument20 pagesLawrance Africa Imagined in The Spanish Renaissance - Henry Thomas LecturejlawranceNo ratings yet
- Prolog Programs Are A Collection of FactsDocument23 pagesProlog Programs Are A Collection of FactsAbni booNo ratings yet
- ResearchDocument38 pagesResearchHelen McClintockNo ratings yet
- Catalog de Produse - BTRDocument62 pagesCatalog de Produse - BTRdaliproiectareNo ratings yet
- Act 1 Almeyda JTLDocument2 pagesAct 1 Almeyda JTLAltairNo ratings yet
- Marik ServitorsDocument20 pagesMarik ServitorsDWNloader100% (2)
- Taylor Swift LyricsDocument2 pagesTaylor Swift LyricsElsie DomeNo ratings yet
- APA-AP - Activity 3-Emerging Ethical and Policy Issues of STSDocument36 pagesAPA-AP - Activity 3-Emerging Ethical and Policy Issues of STSJOHN MARTIN APA-APNo ratings yet
- ContinueDocument3 pagesContinueJúÑi ØrNo ratings yet
- Arlon 38NDocument4 pagesArlon 38NRavindrakumar ParvathiniNo ratings yet
- 501-453 Universal Cable GalndsDocument1 page501-453 Universal Cable Galndsmeribout adelNo ratings yet
- Group 24 ReportDocument48 pagesGroup 24 ReportVishwanath GroupsNo ratings yet
- Glass Configurator Datasheet 2023 03 27Document1 pageGlass Configurator Datasheet 2023 03 27Satrio PrakosoNo ratings yet
- Ethical Considerations Dissertation ExampleDocument7 pagesEthical Considerations Dissertation ExampleBuyPapersOnlineCanada100% (1)
- TakeawayDocument6 pagesTakeawayWilman VasquezNo ratings yet
- Chapter 1 3 THESISDocument64 pagesChapter 1 3 THESISLyka Cartagena-Pandili Manalo-CabelloNo ratings yet
- Drama Bahasa InggrisDocument24 pagesDrama Bahasa InggrisCahyo Yang bahagiaNo ratings yet
- Indias Gold Market Reform and GrowthDocument149 pagesIndias Gold Market Reform and GrowthtmeygmvzjfnkqcwhgpNo ratings yet
- DAPA Teams Spider DiagramDocument2 pagesDAPA Teams Spider DiagramEduardoJaimeNo ratings yet
- Omni-Directional Mobile Robot With Mecanum WheelDocument6 pagesOmni-Directional Mobile Robot With Mecanum WheelTRUNG QUOC LENo ratings yet