Download as pdf or txt
You might also like
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5823)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1093)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (852)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (590)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (898)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (541)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (349)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (823)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (122)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (403)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (74)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- Feminism in IrDocument55 pagesFeminism in IrudayNo ratings yet
- Potfolio Group 1 - Right To EducationDocument33 pagesPotfolio Group 1 - Right To EducationudayNo ratings yet
- Amity School of Engineering & Technology: Submitted To: Submitted byDocument32 pagesAmity School of Engineering & Technology: Submitted To: Submitted byudayNo ratings yet
- Amity School of Engineering & Technology: Submitted To: Submitted byDocument25 pagesAmity School of Engineering & Technology: Submitted To: Submitted byudayNo ratings yet
- Module 5Document82 pagesModule 5udayNo ratings yet
- Reflexive Verbs in SpanishDocument2 pagesReflexive Verbs in SpanishudayNo ratings yet
- Amity University, Uttar Pradesh Department of Electronic &communicationDocument20 pagesAmity University, Uttar Pradesh Department of Electronic &communicationudayNo ratings yet
- Module 2Document67 pagesModule 2udayNo ratings yet
- Module 4Document66 pagesModule 4udayNo ratings yet
- Module 3Document79 pagesModule 3udayNo ratings yet
- Practice Problem 2Document9 pagesPractice Problem 2udayNo ratings yet
- Advanced Simulation Lab: Amity University Noida, Uttar PradeshDocument50 pagesAdvanced Simulation Lab: Amity University Noida, Uttar PradeshudayNo ratings yet
- Module 1Document94 pagesModule 1udayNo ratings yet
- Verilog EXPERIMENT 4Document8 pagesVerilog EXPERIMENT 4udayNo ratings yet
- Verilog EXPERIMENT 2Document11 pagesVerilog EXPERIMENT 2udayNo ratings yet
- Lab FormatDocument1 pageLab FormatudayNo ratings yet
- Assignment 1: Aim: Make A Basic Labview Vi Using Booleans To Compare Some Numbers To Identify TheDocument3 pagesAssignment 1: Aim: Make A Basic Labview Vi Using Booleans To Compare Some Numbers To Identify TheudayNo ratings yet
- NewSyllabus 1910202071612817Document3 pagesNewSyllabus 1910202071612817udayNo ratings yet
- Chapter-1: 1.1. Serial Data TransmissionDocument44 pagesChapter-1: 1.1. Serial Data TransmissionudayNo ratings yet
- Aim:-Verilog Implementation of 8 To 3 PriorityDocument5 pagesAim:-Verilog Implementation of 8 To 3 PriorityudayNo ratings yet
- EXPERIMENT 6 VerilogDocument5 pagesEXPERIMENT 6 VerilogudayNo ratings yet
- Verilog (About Tool and Steps)Document4 pagesVerilog (About Tool and Steps)udayNo ratings yet
- Experiment 10: Aim:-Verilog Implementation of Clock Divider Important Concepts/TheoryDocument6 pagesExperiment 10: Aim:-Verilog Implementation of Clock Divider Important Concepts/TheoryudayNo ratings yet
- EXPERIMENT 8 VerilogDocument6 pagesEXPERIMENT 8 VerilogudayNo ratings yet
- Table 1: Truth Table of An ALUDocument5 pagesTable 1: Truth Table of An ALUudayNo ratings yet
- Experiment 12: Aim:-Software UsedDocument8 pagesExperiment 12: Aim:-Software UsedudayNo ratings yet
- EXPERIMENT 5 VerilogDocument7 pagesEXPERIMENT 5 VerilogudayNo ratings yet
- Experiment 12: Aim: - Important Concepts/TheoryDocument8 pagesExperiment 12: Aim: - Important Concepts/TheoryudayNo ratings yet
- Open Ended Experiment Lab File Verilog Programming (Ece 423) Aim: To Implement Verilog HDL Code For UartDocument3 pagesOpen Ended Experiment Lab File Verilog Programming (Ece 423) Aim: To Implement Verilog HDL Code For UartudayNo ratings yet
- Experiment 10: Aim:-Verilog Implementation of Clock Divider Software UsedDocument6 pagesExperiment 10: Aim:-Verilog Implementation of Clock Divider Software UsedudayNo ratings yet
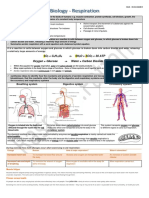
- 6O + C H O - 6H O + 6co + 38 ATP Oxygen + Glucose - Water + Carbon Dioxide + EnergyDocument2 pages6O + C H O - 6H O + 6co + 38 ATP Oxygen + Glucose - Water + Carbon Dioxide + EnergyRacha Abdel Fattah MetwallyNo ratings yet
- Lecture 5Document10 pagesLecture 5Smasher AustineNo ratings yet
- Protein Structure PredictionDocument52 pagesProtein Structure PredictionMudit MisraNo ratings yet
- Biochemistry 1st Edition Miesfeld Test BankDocument36 pagesBiochemistry 1st Edition Miesfeld Test Bankloganhatfieldtkngrzmwcf100% (32)
- Red Cell Membrane Structure 2Document16 pagesRed Cell Membrane Structure 2MONo ratings yet
- Illustrated DIY CRISPR - Freeze Dried DH5a Liquid PlasmidsDocument19 pagesIllustrated DIY CRISPR - Freeze Dried DH5a Liquid PlasmidsKevin LimNo ratings yet
- Science10 Q4 Week-3-BiomoleculesDocument7 pagesScience10 Q4 Week-3-BiomoleculesMaskter ArcheryNo ratings yet
- Oc Amino Acids Peptides and ProteinsDocument11 pagesOc Amino Acids Peptides and ProteinsNhung Lý NgọcNo ratings yet
- Grade 5 Spelling Challenge 3Document2 pagesGrade 5 Spelling Challenge 3TUNG DUONG HANo ratings yet
- Chapterwise Past Papers of MDCATDocument110 pagesChapterwise Past Papers of MDCATKaru malNo ratings yet
- Regents Biology Homework Packet Unit 5: Energy in A Cell Photosynthesis & Cellular RespirationDocument15 pagesRegents Biology Homework Packet Unit 5: Energy in A Cell Photosynthesis & Cellular RespirationHakan Alkan0% (1)
- LUTHFIDocument1 pageLUTHFIbudy raharjaNo ratings yet
- Lab Manual FinalDocument28 pagesLab Manual FinalErwin ManipolNo ratings yet
- Biochemical Functions of LiverDocument94 pagesBiochemical Functions of LiverMi PatelNo ratings yet
- Biology 11th 14 - 01 - 2024 - 001525Document9 pagesBiology 11th 14 - 01 - 2024 - 001525zabiu8926No ratings yet
- Palmit072012slr PDFDocument22 pagesPalmit072012slr PDFGalina TodorovaNo ratings yet
- Ons Oncc Chemotherapy Immunotherapy Certificate Trial New Update Material Guide With Questions and ADocument19 pagesOns Oncc Chemotherapy Immunotherapy Certificate Trial New Update Material Guide With Questions and Amary011danielNo ratings yet
- BOOK TRANS MegakaryopoiesisDocument3 pagesBOOK TRANS MegakaryopoiesisPaul LesterNo ratings yet
- Basic and Applied Cell Physiology: Reference BooksDocument28 pagesBasic and Applied Cell Physiology: Reference Booksjamila ilNo ratings yet
- 2 - CED Reading Alignment Biology in Focus 2nd Edition 2EDocument5 pages2 - CED Reading Alignment Biology in Focus 2nd Edition 2EDani HussainNo ratings yet
- Protein StructuresDocument10 pagesProtein StructuresSebastian RodriguezNo ratings yet
- Fibrous Protein or ScleroproteinDocument4 pagesFibrous Protein or ScleroproteinMia DimagibaNo ratings yet
- Genes AssignmentDocument4 pagesGenes AssignmentKyle Hilary MatundingNo ratings yet
- Lab ManualDocument10 pagesLab ManualozlemNo ratings yet
- Transcription and RNA ProcessingDocument38 pagesTranscription and RNA ProcessingRishi Kumar100% (1)
- Pharmacodynamics Lecture 2Document30 pagesPharmacodynamics Lecture 2Vignesh ReddyNo ratings yet
- VCE Biology Unit 3.1 - Respiration - Main Stages in and Sites of Cellular RespirationDocument6 pagesVCE Biology Unit 3.1 - Respiration - Main Stages in and Sites of Cellular Respirationehung01No ratings yet
- Elsevier's Integrated Review Genetics With STUDENT CONSULT Online Access, 2e (Saunders) (2011)Document270 pagesElsevier's Integrated Review Genetics With STUDENT CONSULT Online Access, 2e (Saunders) (2011)abeer.amerNo ratings yet
- Lessonia-Fucoreverse BrochureDocument5 pagesLessonia-Fucoreverse BrochureElaineSilvestriniNo ratings yet
- Pglo Lab ReportDocument4 pagesPglo Lab Reportapi-345256671No ratings yet