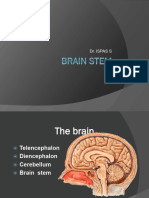
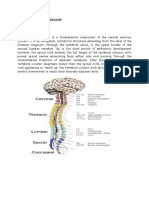
Download as docx, pdf, or txt
You might also like
- ASV RC100 Service ManualDocument89 pagesASV RC100 Service Manualpistol pete100% (1)
- (Robert Mailloux) Electronically Scanned ArraysDocument92 pages(Robert Mailloux) Electronically Scanned ArraysManikanta LalkotaNo ratings yet
- Anatomy SpineDocument6 pagesAnatomy SpineKris AdiNo ratings yet
- Gross Anatomy Draft For PBLDocument10 pagesGross Anatomy Draft For PBLadaumedNo ratings yet
- NeuroembryologyDocument40 pagesNeuroembryologyTaonga jean ChilindaNo ratings yet
- DEVELOPMENT NERVOUS SYSTEM Brain Face PDFDocument15 pagesDEVELOPMENT NERVOUS SYSTEM Brain Face PDFChidera EmmanuelNo ratings yet
- Brainstem (Old Testament)Document19 pagesBrainstem (Old Testament)marizamoraNo ratings yet
- Lesson 3.Document15 pagesLesson 3.Maedel Rose EsguerraNo ratings yet
- Development of Rhombencephalon and MesencephalonDocument27 pagesDevelopment of Rhombencephalon and MesencephalonRafique AhmedNo ratings yet
- Development of the neural tissue topic 4Document4 pagesDevelopment of the neural tissue topic 4omarmamluky254No ratings yet
- Development of Brain:: Spinal CordDocument3 pagesDevelopment of Brain:: Spinal Cordhaseeb ShafaatNo ratings yet
- Cerebellum TextDocument7 pagesCerebellum TextJean Pierre Chastre LuzaNo ratings yet
- EmbryologyDocument92 pagesEmbryologyLoveena SteadmanNo ratings yet
- Cerebellum and HypothalamusDocument20 pagesCerebellum and HypothalamusSaviourNo ratings yet
- Brain StemDocument47 pagesBrain StemSorina IspasNo ratings yet
- The CerebellumDocument5 pagesThe CerebellumAndrew BarryNo ratings yet
- Lecture - Slides-01 - 06 - Medial Surface of The BrainDocument6 pagesLecture - Slides-01 - 06 - Medial Surface of The BrainMaria Eduarda VianaNo ratings yet
- Spinal CordDocument9 pagesSpinal Cordpromisearinzechi4No ratings yet
- Neuroanatomy, Cerebellar Dysfunction: Gross AnatomyDocument5 pagesNeuroanatomy, Cerebellar Dysfunction: Gross AnatomyANISA RIFKA RIDHONo ratings yet
- Brain AnatomyDocument9 pagesBrain AnatomyFikertemariam DessalegnNo ratings yet
- The Descending Tracts of The Spinal CordDocument2 pagesThe Descending Tracts of The Spinal Cordalmastar officeNo ratings yet
- The BrainDocument31 pagesThe BrainHusseinNo ratings yet
- Alberta Stroke Program Early CT Score - 1Document4 pagesAlberta Stroke Program Early CT Score - 1Ardina Nur PramudhitaNo ratings yet
- AD 5 Vertebral AnatomyDocument37 pagesAD 5 Vertebral AnatomyLintang ZerlinaNo ratings yet
- Department of Anatomy College of Medicine University of Ibadan Neuroanatomy Dissection of The BrainDocument22 pagesDepartment of Anatomy College of Medicine University of Ibadan Neuroanatomy Dissection of The BrainAbdulateef AdebisiNo ratings yet
- Anat CNS QBDocument22 pagesAnat CNS QBSATVIK RAMNo ratings yet
- LAB EXERCISE 4 - 4 MM Frog EmbryoDocument7 pagesLAB EXERCISE 4 - 4 MM Frog EmbryoGerald Angelo DeguinioNo ratings yet
- Karen Flores Assenet Pascual Paloma Priscila Tania Sanchez Aime Murillo Alberto MorenoDocument11 pagesKaren Flores Assenet Pascual Paloma Priscila Tania Sanchez Aime Murillo Alberto MorenoTania Berenice Sanchez MuroNo ratings yet
- Aprimaion Angulo PontocerebelosoDocument64 pagesAprimaion Angulo PontocerebelosoLuis Carlos Avellaneda CurchoNo ratings yet
- The CerebellumDocument19 pagesThe Cerebellumalecsandra96No ratings yet
- Development of The Central Nervous System: Consists of The Brain and Spinal Cord Derived From The Neural TubeDocument42 pagesDevelopment of The Central Nervous System: Consists of The Brain and Spinal Cord Derived From The Neural TubeZyra PerolNo ratings yet
- 1.1.6 NeuroanatomyDocument10 pages1.1.6 NeuroanatomyZayan SyedNo ratings yet
- Lecture 3 Anatomy 4 Spring 2023 PDFDocument18 pagesLecture 3 Anatomy 4 Spring 2023 PDFAhmed NaeemNo ratings yet
- BRAINSTEMDocument17 pagesBRAINSTEMaroosadilbar27No ratings yet
- Brain Stem: Rabbia Yousaf (Pharm-D) Rashid Latif College of PharmacyDocument30 pagesBrain Stem: Rabbia Yousaf (Pharm-D) Rashid Latif College of PharmacyShafaqat Ghani Shafaqat GhaniNo ratings yet
- (Journal of Neurosurgery) Microsurgical Anatomy of The Veins of The Posterior FossaDocument43 pages(Journal of Neurosurgery) Microsurgical Anatomy of The Veins of The Posterior FossaCan EkerNo ratings yet
- Embryology of The CNS 2022Document130 pagesEmbryology of The CNS 2022isicheipraise3No ratings yet
- Overview: Functions of The CerebellumDocument17 pagesOverview: Functions of The CerebellumMohd Fitri ShahruddinNo ratings yet
- Embryology Summary of Development Part 1 - 2019 - CNS, Axial SK, SK Muscle, SkinDocument9 pagesEmbryology Summary of Development Part 1 - 2019 - CNS, Axial SK, SK Muscle, Skindr.mumtaz09No ratings yet
- 02 - CNS DevelopmentDocument3 pages02 - CNS DevelopmentCemre KuzeyNo ratings yet
- Embriologi Brain and SpineDocument18 pagesEmbriologi Brain and SpineJunus EufrataNo ratings yet
- 1 - Spinal Cord NotesDocument7 pages1 - Spinal Cord NotesOceanChan QdNo ratings yet
- 13 Anatomy of The Nervous SystemDocument95 pages13 Anatomy of The Nervous SystemolivNo ratings yet
- Neuroanatomy 1Document65 pagesNeuroanatomy 1maNo ratings yet
- CNS - Embryology MorningDocument34 pagesCNS - Embryology MorningLugembe MasolaNo ratings yet
- 1 My Trigeminal NerveDocument23 pages1 My Trigeminal NerveDan 04No ratings yet
- Embryology and Anatomy: Spine/Spinal CordDocument5 pagesEmbryology and Anatomy: Spine/Spinal CordFernandoo CunhaNo ratings yet
- Growth and Development of Cranial Base: SeminarDocument34 pagesGrowth and Development of Cranial Base: SeminarMNSNo ratings yet
- Anatomy and Physiology The Spinal CordDocument11 pagesAnatomy and Physiology The Spinal CordKatrina CaveNo ratings yet
- Centralnervoussystem CNSDocument19 pagesCentralnervoussystem CNSÂhmęd ĐoolaNo ratings yet
- Cranial BaseDocument25 pagesCranial BasesealovesriverNo ratings yet
- Embryology-Development of Central Nervous SystemDocument28 pagesEmbryology-Development of Central Nervous Systemtenshi315No ratings yet
- 10 CerebellumDocument26 pages10 CerebellumDarpan KumariNo ratings yet
- Ant 325materialDocument13 pagesAnt 325materialDestiny BlissNo ratings yet
- Crossman-book-summary-fullDocument53 pagesCrossman-book-summary-fullDaniele TomassiNo ratings yet
- ЦНС - 2019Document50 pagesЦНС - 2019bekafop813No ratings yet
- Thalamus NotesDocument11 pagesThalamus NotesTanvir AhmedNo ratings yet
- Brainstem: Medulla OblongataDocument130 pagesBrainstem: Medulla OblongataBhawana BhatodiyaNo ratings yet
- Chapter IIDocument29 pagesChapter IIKurnia pralisaNo ratings yet
- Thespinalcord: A Review of Functional NeuroanatomyDocument18 pagesThespinalcord: A Review of Functional NeuroanatomyasfwegereNo ratings yet
- 13 Anatomy of The Nervous SystemDocument78 pages13 Anatomy of The Nervous SystemolivNo ratings yet
- VTK File FormatsDocument19 pagesVTK File FormatshamishlewisNo ratings yet
- Test Bank For Finite Mathematics and Its Applications 12Th by Goldstein Schneider Siegel and Hair Isbn 0134437764 9780134437767 Full Chapter PDFDocument36 pagesTest Bank For Finite Mathematics and Its Applications 12Th by Goldstein Schneider Siegel and Hair Isbn 0134437764 9780134437767 Full Chapter PDFjeremy.curiel650100% (18)
- Fundamentals of Computer GraphicsDocument58 pagesFundamentals of Computer GraphicsISHAAN TAKKARNo ratings yet
- Head LukasDocument43 pagesHead LukasSergent Khaos100% (1)
- LFCS Domains Competencies V2.16 PDFDocument7 pagesLFCS Domains Competencies V2.16 PDFKeny Oscar Cortes GonzalezNo ratings yet
- Micro X Ray Fluorescence Spectroscopy First Edition PDFDocument31 pagesMicro X Ray Fluorescence Spectroscopy First Edition PDFAlberto Núñez CardezoNo ratings yet
- H3D Manual - Complete ABDZP0GADocument63 pagesH3D Manual - Complete ABDZP0GAMiladShahabiNo ratings yet
- Residential Multi Split CatalogueDocument51 pagesResidential Multi Split Cataloguengocdhxd92No ratings yet
- Textbook Engineering Economics Financial Decision Making For Engineers 5Th Edition Niall Fraser Ebook All Chapter PDFDocument33 pagesTextbook Engineering Economics Financial Decision Making For Engineers 5Th Edition Niall Fraser Ebook All Chapter PDFjennifer.bias411100% (15)
- DC Protection Relay PrinciplesDocument28 pagesDC Protection Relay Principlesrvim0002100% (1)
- ML Ass 4Document7 pagesML Ass 4Santhosh Kumar PNo ratings yet
- Maia Grammar Essentials PDFDocument176 pagesMaia Grammar Essentials PDFRauf Mammadov100% (1)
- Hendrickson Assignment4Document11 pagesHendrickson Assignment4Angel Nicole SorianoNo ratings yet
- Classified Paper 2 Part 1Document408 pagesClassified Paper 2 Part 1LIN YAN100% (1)
- KSB PUMPS Co. Ama DrainerDocument2 pagesKSB PUMPS Co. Ama DrainerHuzaifa KhanNo ratings yet
- Devinfo E6350v2Document4 pagesDevinfo E6350v2Safi Ud Din KhanNo ratings yet
- Not Recommended For New Design: Parallel Port Button HolderDocument2 pagesNot Recommended For New Design: Parallel Port Button HolderSaasiNo ratings yet
- Mass Balance Calculations - March 22 - FermentationDocument13 pagesMass Balance Calculations - March 22 - FermentationRj JunsayNo ratings yet
- SURveying IIDocument159 pagesSURveying IIpieterNo ratings yet
- Syntax: The Sentence Patterns of Language: 01:615:201 Introduction To Linguistic TheoryDocument54 pagesSyntax: The Sentence Patterns of Language: 01:615:201 Introduction To Linguistic TheoryAmalia AsokawatiNo ratings yet
- Ce 023 Module 5 and 6Document9 pagesCe 023 Module 5 and 6George YanelaNo ratings yet
- Sound Short Answer Based Questions: S.N. Column A (Frequency)Document6 pagesSound Short Answer Based Questions: S.N. Column A (Frequency)Madhukar BhagatNo ratings yet
- O o o o o o o o o o oDocument17 pagesO o o o o o o o o o oshukkurNo ratings yet
- Math 32 - 06 - Bayes Rule (Examples)Document7 pagesMath 32 - 06 - Bayes Rule (Examples)Celine ShawNo ratings yet
- ElectrolightfinalDocument12 pagesElectrolightfinalMigs GisonNo ratings yet
- Tool ManagementDocument6 pagesTool ManagementRoshan RamnaniNo ratings yet
- Chart Work - OowDocument159 pagesChart Work - OowAboody AL-ghamdyNo ratings yet
- Weidmuller SAI PIDocument32 pagesWeidmuller SAI PIHrvoje HorvatNo ratings yet