
Non-Homologous End Joining
Non-Homologous End Joining
You might also like
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5820)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1093)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (845)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (590)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (898)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (540)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (349)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (822)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (122)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (401)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (74)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- (Methods in Enzymology 463) Richard R. Burgess and Murray P. Deutscher (Eds.) - Guide To Protein Purification, 2nd Edition-Academic Press (2009)Document853 pages(Methods in Enzymology 463) Richard R. Burgess and Murray P. Deutscher (Eds.) - Guide To Protein Purification, 2nd Edition-Academic Press (2009)Josephine Tan100% (1)
- Integrin: Integrin Alphavbeta3 Extracellular DomainsDocument15 pagesIntegrin: Integrin Alphavbeta3 Extracellular Domainsfelipe smithNo ratings yet
- Upload 1 Document To Download: Incendiaries Advanced Improvised ExplosivesDocument3 pagesUpload 1 Document To Download: Incendiaries Advanced Improvised Explosivesfelipe smithNo ratings yet
- Teste 1Document44 pagesTeste 1felipe smithNo ratings yet
- Home Search Your Library: Spoti - 402,030 LikesDocument1 pageHome Search Your Library: Spoti - 402,030 Likesfelipe smithNo ratings yet
- Neural Cell Adhesion MoleculeDocument7 pagesNeural Cell Adhesion Moleculefelipe smithNo ratings yet
- Sustainable Steel Construction20160812 385 1yxj1ey With Cover Page v2Document19 pagesSustainable Steel Construction20160812 385 1yxj1ey With Cover Page v2felipe smithNo ratings yet
- Von Willebrand Factor (VWF)Document9 pagesVon Willebrand Factor (VWF)felipe smithNo ratings yet
- Glycoprotein: Glycoproteins Are Proteins Which Contain Oligosaccharide ChainsDocument8 pagesGlycoprotein: Glycoproteins Are Proteins Which Contain Oligosaccharide Chainsfelipe smithNo ratings yet
- Cystic Fibrosis Transmembrane Conductance Regulator (CFTR) Is ADocument17 pagesCystic Fibrosis Transmembrane Conductance Regulator (CFTR) Is Afelipe smithNo ratings yet
- Thrombin: Thrombin (EC 3.4.21.5 (Https://enzDocument17 pagesThrombin: Thrombin (EC 3.4.21.5 (Https://enzfelipe smithNo ratings yet
- Protein Z-Related Protease InhibitorDocument2 pagesProtein Z-Related Protease Inhibitorfelipe smithNo ratings yet
- Protein S (Also Known As S-Protein)Document10 pagesProtein S (Also Known As S-Protein)felipe smithNo ratings yet
- C3-Convertase: C3 Convertase (C4bC2b, Formerly C4bC2a) Belongs To Family ofDocument5 pagesC3-Convertase: C3 Convertase (C4bC2b, Formerly C4bC2a) Belongs To Family offelipe smithNo ratings yet
- DNA Polymerase Lambda: FunctionDocument8 pagesDNA Polymerase Lambda: Functionfelipe smithNo ratings yet
- Serum Amyloid P Component: in AmyloidosisDocument5 pagesSerum Amyloid P Component: in Amyloidosisfelipe smithNo ratings yet
- Cadherin: Cadherins (Named For "Calcium-DependentDocument10 pagesCadherin: Cadherins (Named For "Calcium-Dependentfelipe smithNo ratings yet
- Protein C: Protein C, Autoprothrombin IIA and Blood Coagulation Factor XIXDocument14 pagesProtein C: Protein C, Autoprothrombin IIA and Blood Coagulation Factor XIXfelipe smithNo ratings yet
- Protein ZDocument3 pagesProtein Zfelipe smithNo ratings yet
- Elastin: Elastin Is A Key Protein of TheDocument12 pagesElastin: Elastin Is A Key Protein of Thefelipe smithNo ratings yet
- Serum Albumin: Serum Albumin, Often Referred To Simply As Blood Albumin, Is AnDocument4 pagesSerum Albumin: Serum Albumin, Often Referred To Simply As Blood Albumin, Is Anfelipe smithNo ratings yet
- Fibronectin: Fibronectin Is A High-MolecularDocument22 pagesFibronectin: Fibronectin Is A High-Molecularfelipe smithNo ratings yet
- Tubulin: Tubulin in Molecular Biology Can Refer Either To The Tubulin ProteinDocument11 pagesTubulin: Tubulin in Molecular Biology Can Refer Either To The Tubulin Proteinfelipe smithNo ratings yet
- Spondin 1: ReferencesDocument9 pagesSpondin 1: Referencesfelipe smithNo ratings yet
- Artemis Is A Protein That in HumansDocument10 pagesArtemis Is A Protein That in Humansfelipe smithNo ratings yet
- DNA Polymerase Mu Is ADocument6 pagesDNA Polymerase Mu Is Afelipe smithNo ratings yet
- Arp2/3 Complex: Arp2/3 Complex Is A Seven-Subunit Protein Complex That Plays ADocument5 pagesArp2/3 Complex: Arp2/3 Complex Is A Seven-Subunit Protein Complex That Plays Afelipe smithNo ratings yet
- KeratinDocument8 pagesKeratinfelipe smithNo ratings yet
- FTSZ Is A Protein Encoded by The FTSZ Gene That Assembles Into A RingDocument8 pagesFTSZ Is A Protein Encoded by The FTSZ Gene That Assembles Into A Ringfelipe smithNo ratings yet
- Bifunctional Polynucleotide Phosphatase/kinase Is An EnzymeDocument10 pagesBifunctional Polynucleotide Phosphatase/kinase Is An Enzymefelipe smithNo ratings yet
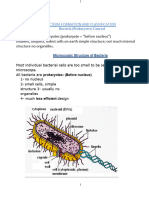
- Bacteria Formation ClassificationDocument9 pagesBacteria Formation Classificationfurqanbaig8No ratings yet
- LFSC Autumn Camp 2023 Marking GuidelinesDocument16 pagesLFSC Autumn Camp 2023 Marking GuidelinesmanotosbongileNo ratings yet
- Hamcus Wiki enDocument17 pagesHamcus Wiki envocew91486No ratings yet
- DNA Repair MechanismDocument4 pagesDNA Repair MechanismIraqi100% (1)
- Biology Project DheekshanyaDocument17 pagesBiology Project DheekshanyaDheekshith KumarNo ratings yet
- Cómo Escribir Un Ensayo A Partir de Una EntrevistaDocument4 pagesCómo Escribir Un Ensayo A Partir de Una Entrevistacjb93mda100% (1)
- Mendelian GeneticsDocument71 pagesMendelian GeneticsUmutFerhatBaşpınarNo ratings yet
- Genetic Algorithms: Aroob Amjad AhmedDocument5 pagesGenetic Algorithms: Aroob Amjad AhmedAroob amjadNo ratings yet
- Historical Development of Cell TheoryDocument31 pagesHistorical Development of Cell TheoryRamona BaculinaoNo ratings yet
- 9th June, 2021 Daily Global Regional Local Rice E-NewsletterDocument56 pages9th June, 2021 Daily Global Regional Local Rice E-NewsletterMujahid AliNo ratings yet
- Anthropology - Test - Series - Chapter WiseDocument7 pagesAnthropology - Test - Series - Chapter Wisedhivya sankarNo ratings yet
- Bonobos Are Only Genetically Similar To HumansDocument2 pagesBonobos Are Only Genetically Similar To HumansAndrea GNo ratings yet
- CHEMISTRY 350 Fall 2023 Examination 1Document6 pagesCHEMISTRY 350 Fall 2023 Examination 1hibadesi00No ratings yet
- Ra Uru Hu ProfileDocument3 pagesRa Uru Hu Profilebob100% (1)
- Microevolution: Micro-Macro - & MegaevolutionDocument2 pagesMicroevolution: Micro-Macro - & MegaevolutionAnupama KNo ratings yet
- Race Ission Ollege: Issues On Human DevelopmentDocument7 pagesRace Ission Ollege: Issues On Human DevelopmentElle Aquino100% (3)
- DNA AnalysisDocument39 pagesDNA AnalysisBio SciencesNo ratings yet
- DNA ReplicationDocument3 pagesDNA ReplicationXyrelle EspielNo ratings yet
- DNA Conformation and Transcription Pag. 145 SupercoilingDocument215 pagesDNA Conformation and Transcription Pag. 145 SupercoilingDiegoNo ratings yet
- Performance of Popcorn Introductions For AgronomicDocument8 pagesPerformance of Popcorn Introductions For AgronomicDANIEL LEONARDO DE LA CRUZ GOMEZNo ratings yet
- Lab MitosisDocument4 pagesLab Mitosisnowlik56100% (7)
- Department of EducationDocument16 pagesDepartment of EducationDannica De La CruzNo ratings yet
- Apraxia of Speech and Grammatical Language Impairment in Children With Autism Procedural Deficit HypothesisDocument6 pagesApraxia of Speech and Grammatical Language Impairment in Children With Autism Procedural Deficit HypothesisEditor IJTSRDNo ratings yet
- Cambridge International AS & A Level: Biology 9700/12Document16 pagesCambridge International AS & A Level: Biology 9700/12Milinda De SilvaNo ratings yet
- Faculty of Psychology Bachelor of Science (Psychology) Curriculum Structure Document Academic YearDocument97 pagesFaculty of Psychology Bachelor of Science (Psychology) Curriculum Structure Document Academic YearKashyapi thakuriaNo ratings yet
- Potential Applications of Molecular Markers in Plant: Mini ReviewDocument3 pagesPotential Applications of Molecular Markers in Plant: Mini ReviewFernando Castro EchavezNo ratings yet
- The Selection Process WorksheetDocument2 pagesThe Selection Process Worksheetapi-256236481No ratings yet
- BasicsDocument18 pagesBasicsBiraj KarmakarNo ratings yet
- Moleculer Variance Identication of Commercial Genotipe Oil Palm (Elaeis Guineensis Jacq.) Based On Random Amplified Polymorpism DNA (RAPD) MarkersDocument4 pagesMoleculer Variance Identication of Commercial Genotipe Oil Palm (Elaeis Guineensis Jacq.) Based On Random Amplified Polymorpism DNA (RAPD) MarkersArnen PasaribuNo ratings yet


































































































Download as pdf or txt
You might also like
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5820)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1093)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (845)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (590)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (898)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (540)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (349)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (822)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (122)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (401)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (74)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- (Methods in Enzymology 463) Richard R. Burgess and Murray P. Deutscher (Eds.) - Guide To Protein Purification, 2nd Edition-Academic Press (2009)Document853 pages(Methods in Enzymology 463) Richard R. Burgess and Murray P. Deutscher (Eds.) - Guide To Protein Purification, 2nd Edition-Academic Press (2009)Josephine Tan100% (1)
- Integrin: Integrin Alphavbeta3 Extracellular DomainsDocument15 pagesIntegrin: Integrin Alphavbeta3 Extracellular Domainsfelipe smithNo ratings yet
- Upload 1 Document To Download: Incendiaries Advanced Improvised ExplosivesDocument3 pagesUpload 1 Document To Download: Incendiaries Advanced Improvised Explosivesfelipe smithNo ratings yet
- Teste 1Document44 pagesTeste 1felipe smithNo ratings yet
- Home Search Your Library: Spoti - 402,030 LikesDocument1 pageHome Search Your Library: Spoti - 402,030 Likesfelipe smithNo ratings yet
- Neural Cell Adhesion MoleculeDocument7 pagesNeural Cell Adhesion Moleculefelipe smithNo ratings yet
- Sustainable Steel Construction20160812 385 1yxj1ey With Cover Page v2Document19 pagesSustainable Steel Construction20160812 385 1yxj1ey With Cover Page v2felipe smithNo ratings yet
- Von Willebrand Factor (VWF)Document9 pagesVon Willebrand Factor (VWF)felipe smithNo ratings yet
- Glycoprotein: Glycoproteins Are Proteins Which Contain Oligosaccharide ChainsDocument8 pagesGlycoprotein: Glycoproteins Are Proteins Which Contain Oligosaccharide Chainsfelipe smithNo ratings yet
- Cystic Fibrosis Transmembrane Conductance Regulator (CFTR) Is ADocument17 pagesCystic Fibrosis Transmembrane Conductance Regulator (CFTR) Is Afelipe smithNo ratings yet
- Thrombin: Thrombin (EC 3.4.21.5 (Https://enzDocument17 pagesThrombin: Thrombin (EC 3.4.21.5 (Https://enzfelipe smithNo ratings yet
- Protein Z-Related Protease InhibitorDocument2 pagesProtein Z-Related Protease Inhibitorfelipe smithNo ratings yet
- Protein S (Also Known As S-Protein)Document10 pagesProtein S (Also Known As S-Protein)felipe smithNo ratings yet
- C3-Convertase: C3 Convertase (C4bC2b, Formerly C4bC2a) Belongs To Family ofDocument5 pagesC3-Convertase: C3 Convertase (C4bC2b, Formerly C4bC2a) Belongs To Family offelipe smithNo ratings yet
- DNA Polymerase Lambda: FunctionDocument8 pagesDNA Polymerase Lambda: Functionfelipe smithNo ratings yet
- Serum Amyloid P Component: in AmyloidosisDocument5 pagesSerum Amyloid P Component: in Amyloidosisfelipe smithNo ratings yet
- Cadherin: Cadherins (Named For "Calcium-DependentDocument10 pagesCadherin: Cadherins (Named For "Calcium-Dependentfelipe smithNo ratings yet
- Protein C: Protein C, Autoprothrombin IIA and Blood Coagulation Factor XIXDocument14 pagesProtein C: Protein C, Autoprothrombin IIA and Blood Coagulation Factor XIXfelipe smithNo ratings yet
- Protein ZDocument3 pagesProtein Zfelipe smithNo ratings yet
- Elastin: Elastin Is A Key Protein of TheDocument12 pagesElastin: Elastin Is A Key Protein of Thefelipe smithNo ratings yet
- Serum Albumin: Serum Albumin, Often Referred To Simply As Blood Albumin, Is AnDocument4 pagesSerum Albumin: Serum Albumin, Often Referred To Simply As Blood Albumin, Is Anfelipe smithNo ratings yet
- Fibronectin: Fibronectin Is A High-MolecularDocument22 pagesFibronectin: Fibronectin Is A High-Molecularfelipe smithNo ratings yet
- Tubulin: Tubulin in Molecular Biology Can Refer Either To The Tubulin ProteinDocument11 pagesTubulin: Tubulin in Molecular Biology Can Refer Either To The Tubulin Proteinfelipe smithNo ratings yet
- Spondin 1: ReferencesDocument9 pagesSpondin 1: Referencesfelipe smithNo ratings yet
- Artemis Is A Protein That in HumansDocument10 pagesArtemis Is A Protein That in Humansfelipe smithNo ratings yet
- DNA Polymerase Mu Is ADocument6 pagesDNA Polymerase Mu Is Afelipe smithNo ratings yet
- Arp2/3 Complex: Arp2/3 Complex Is A Seven-Subunit Protein Complex That Plays ADocument5 pagesArp2/3 Complex: Arp2/3 Complex Is A Seven-Subunit Protein Complex That Plays Afelipe smithNo ratings yet
- KeratinDocument8 pagesKeratinfelipe smithNo ratings yet
- FTSZ Is A Protein Encoded by The FTSZ Gene That Assembles Into A RingDocument8 pagesFTSZ Is A Protein Encoded by The FTSZ Gene That Assembles Into A Ringfelipe smithNo ratings yet
- Bifunctional Polynucleotide Phosphatase/kinase Is An EnzymeDocument10 pagesBifunctional Polynucleotide Phosphatase/kinase Is An Enzymefelipe smithNo ratings yet
- Bacteria Formation ClassificationDocument9 pagesBacteria Formation Classificationfurqanbaig8No ratings yet
- LFSC Autumn Camp 2023 Marking GuidelinesDocument16 pagesLFSC Autumn Camp 2023 Marking GuidelinesmanotosbongileNo ratings yet
- Hamcus Wiki enDocument17 pagesHamcus Wiki envocew91486No ratings yet
- DNA Repair MechanismDocument4 pagesDNA Repair MechanismIraqi100% (1)
- Biology Project DheekshanyaDocument17 pagesBiology Project DheekshanyaDheekshith KumarNo ratings yet
- Cómo Escribir Un Ensayo A Partir de Una EntrevistaDocument4 pagesCómo Escribir Un Ensayo A Partir de Una Entrevistacjb93mda100% (1)
- Mendelian GeneticsDocument71 pagesMendelian GeneticsUmutFerhatBaşpınarNo ratings yet
- Genetic Algorithms: Aroob Amjad AhmedDocument5 pagesGenetic Algorithms: Aroob Amjad AhmedAroob amjadNo ratings yet
- Historical Development of Cell TheoryDocument31 pagesHistorical Development of Cell TheoryRamona BaculinaoNo ratings yet
- 9th June, 2021 Daily Global Regional Local Rice E-NewsletterDocument56 pages9th June, 2021 Daily Global Regional Local Rice E-NewsletterMujahid AliNo ratings yet
- Anthropology - Test - Series - Chapter WiseDocument7 pagesAnthropology - Test - Series - Chapter Wisedhivya sankarNo ratings yet
- Bonobos Are Only Genetically Similar To HumansDocument2 pagesBonobos Are Only Genetically Similar To HumansAndrea GNo ratings yet
- CHEMISTRY 350 Fall 2023 Examination 1Document6 pagesCHEMISTRY 350 Fall 2023 Examination 1hibadesi00No ratings yet
- Ra Uru Hu ProfileDocument3 pagesRa Uru Hu Profilebob100% (1)
- Microevolution: Micro-Macro - & MegaevolutionDocument2 pagesMicroevolution: Micro-Macro - & MegaevolutionAnupama KNo ratings yet
- Race Ission Ollege: Issues On Human DevelopmentDocument7 pagesRace Ission Ollege: Issues On Human DevelopmentElle Aquino100% (3)
- DNA AnalysisDocument39 pagesDNA AnalysisBio SciencesNo ratings yet
- DNA ReplicationDocument3 pagesDNA ReplicationXyrelle EspielNo ratings yet
- DNA Conformation and Transcription Pag. 145 SupercoilingDocument215 pagesDNA Conformation and Transcription Pag. 145 SupercoilingDiegoNo ratings yet
- Performance of Popcorn Introductions For AgronomicDocument8 pagesPerformance of Popcorn Introductions For AgronomicDANIEL LEONARDO DE LA CRUZ GOMEZNo ratings yet
- Lab MitosisDocument4 pagesLab Mitosisnowlik56100% (7)
- Department of EducationDocument16 pagesDepartment of EducationDannica De La CruzNo ratings yet
- Apraxia of Speech and Grammatical Language Impairment in Children With Autism Procedural Deficit HypothesisDocument6 pagesApraxia of Speech and Grammatical Language Impairment in Children With Autism Procedural Deficit HypothesisEditor IJTSRDNo ratings yet
- Cambridge International AS & A Level: Biology 9700/12Document16 pagesCambridge International AS & A Level: Biology 9700/12Milinda De SilvaNo ratings yet
- Faculty of Psychology Bachelor of Science (Psychology) Curriculum Structure Document Academic YearDocument97 pagesFaculty of Psychology Bachelor of Science (Psychology) Curriculum Structure Document Academic YearKashyapi thakuriaNo ratings yet
- Potential Applications of Molecular Markers in Plant: Mini ReviewDocument3 pagesPotential Applications of Molecular Markers in Plant: Mini ReviewFernando Castro EchavezNo ratings yet
- The Selection Process WorksheetDocument2 pagesThe Selection Process Worksheetapi-256236481No ratings yet
- BasicsDocument18 pagesBasicsBiraj KarmakarNo ratings yet
- Moleculer Variance Identication of Commercial Genotipe Oil Palm (Elaeis Guineensis Jacq.) Based On Random Amplified Polymorpism DNA (RAPD) MarkersDocument4 pagesMoleculer Variance Identication of Commercial Genotipe Oil Palm (Elaeis Guineensis Jacq.) Based On Random Amplified Polymorpism DNA (RAPD) MarkersArnen PasaribuNo ratings yet