Download as docx, pdf, or txt
You might also like
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5825)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1093)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (852)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (590)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (903)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (541)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (349)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (823)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (122)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (403)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (74)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- BG2109-T1 TeoDocument2 pagesBG2109-T1 TeoLam Kwang YangNo ratings yet
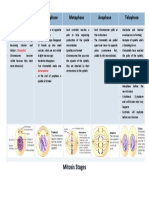
- Early Prophase Late Prophase Metaphase Anaphase Telophase: Mitosis StagesDocument1 pageEarly Prophase Late Prophase Metaphase Anaphase Telophase: Mitosis StagesYasmine HadiastrianiNo ratings yet
- Student Exploration: Cell Division: Isabella Jacquin Rivas November 24, 2022Document5 pagesStudent Exploration: Cell Division: Isabella Jacquin Rivas November 24, 2022。Poki 。No ratings yet
- Parts of The Cell Worksheet - BiochemistryDocument3 pagesParts of The Cell Worksheet - BiochemistryAnonymous giver100% (1)
- Ap Cell Tour 1 ProteinDocument28 pagesAp Cell Tour 1 Proteinapi-235744933100% (1)
- Biology CH 7 Practice TestDocument6 pagesBiology CH 7 Practice Testvaleria100% (2)
- CLS Aipmt 16 17 XIII Bot Study Package 2 SET 1 Chapter 6Document37 pagesCLS Aipmt 16 17 XIII Bot Study Package 2 SET 1 Chapter 6prism1702100% (2)
- CMB PPT 3 Animal Cell Borja Jayven CDocument12 pagesCMB PPT 3 Animal Cell Borja Jayven CJayven BorjaNo ratings yet
- BSC 250 Chapter 3 Blank NotesDocument10 pagesBSC 250 Chapter 3 Blank NotessarahreyNo ratings yet
- Class 9th MCQsDocument8 pagesClass 9th MCQsAmit PratapNo ratings yet
- Human Genetics Concepts and Applications 11th Edition Ricki Lewis Solutions ManualDocument24 pagesHuman Genetics Concepts and Applications 11th Edition Ricki Lewis Solutions ManualMichaelMorgandzkam100% (34)
- Chapter 2 Foundations: The Cell: Human Anatomy, 7e (Martini/Timmons/Tallitsch)Document7 pagesChapter 2 Foundations: The Cell: Human Anatomy, 7e (Martini/Timmons/Tallitsch)admin212No ratings yet
- IB Released Questions On 01 Cell BiologyDocument50 pagesIB Released Questions On 01 Cell BiologyZuzanna BaligaNo ratings yet
- The Cytoskeleton: Cell Biology 1st SemesterDocument22 pagesThe Cytoskeleton: Cell Biology 1st SemesterJason CenaNo ratings yet
- Ultra Structure of ChromosomeDocument9 pagesUltra Structure of ChromosomeJigarS.MehtaNo ratings yet
- Prokariotik Dan Eukariotik 2Document22 pagesProkariotik Dan Eukariotik 2hanumNo ratings yet
- St. Karen's Secondary School, Patna Subject-Biology Class - 8 Chapter - The Cell Answer Key of (Assignment - 3)Document3 pagesSt. Karen's Secondary School, Patna Subject-Biology Class - 8 Chapter - The Cell Answer Key of (Assignment - 3)Harshit RajNo ratings yet
- St. Paul College of Ilocos SurDocument6 pagesSt. Paul College of Ilocos SurRica machells DaydaNo ratings yet
- Plant Cells - A Biology EssayDocument1 pagePlant Cells - A Biology EssayMerNo ratings yet
- pr7IfQFJTnaR deKnLv7lhrDocument7 pagespr7IfQFJTnaR deKnLv7lhrSamarthsinh VaghelaNo ratings yet
- Chapter 3 Cell Division SB015Document59 pagesChapter 3 Cell Division SB015Muhammad IzuanNo ratings yet
- Cell Function StructureDocument3 pagesCell Function StructureVeden Dumogho100% (1)
- Euglena BioDocument30 pagesEuglena BioMaddie KeatingNo ratings yet
- Are Plant & Animal Cells The Same or Different?Document18 pagesAre Plant & Animal Cells The Same or Different?Josa Camille BungayNo ratings yet
- Biotech 3 Cell DivisionDocument4 pagesBiotech 3 Cell DivisionClinton YmbongNo ratings yet
- NotesDocument49 pagesNotesdeepNo ratings yet
- 01-Cell DivisionDocument21 pages01-Cell DivisionSENGUPTA CHANDANNo ratings yet
- Cell Structure 3Document30 pagesCell Structure 3Rohan PaneruNo ratings yet
- Notes Plant Cell 21Document2 pagesNotes Plant Cell 21Samantha JerezNo ratings yet
- Granular Endoplasmic ReticulumDocument2 pagesGranular Endoplasmic ReticulumSetumil PowerNo ratings yet