Download as pdf or txt
You might also like
- JP 2-0 Joint IntelligenceDocument150 pagesJP 2-0 Joint Intelligenceshakes21778No ratings yet
- (Rafael Lira, Alejandro Casas, José Blancas (Eds. PDFDocument562 pages(Rafael Lira, Alejandro Casas, José Blancas (Eds. PDFJOSE FRANCISCO FRANCO NAVIANo ratings yet
- Earth Microbial Co-Occurrence Network Reveals Interconnection Pattern Across MicrobiomesDocument12 pagesEarth Microbial Co-Occurrence Network Reveals Interconnection Pattern Across Microbiomeshan zhouNo ratings yet
- Unifying The Functional Diversity in Natural and CDocument15 pagesUnifying The Functional Diversity in Natural and CGustavo Andres Fuentes RodriguezNo ratings yet
- Genes 13 02280 v3Document34 pagesGenes 13 02280 v3João Vitor Aires TeixeiraNo ratings yet
- Types of Vegetables Shape Composition, Diversity, and Co-Occurrence Networks of Soil Bacteria and Fungi in Karst Areas of Southwest ChinaDocument14 pagesTypes of Vegetables Shape Composition, Diversity, and Co-Occurrence Networks of Soil Bacteria and Fungi in Karst Areas of Southwest ChinaAbel Abelito Jimenez Jimenez Jr.No ratings yet
- The Microbial Dark Matter and Wanted List in WorldDocument16 pagesThe Microbial Dark Matter and Wanted List in WorldmizbaNo ratings yet
- Oportunidades e Desafios Do Uso de Dados MetagenômicosDocument14 pagesOportunidades e Desafios Do Uso de Dados MetagenômicosVictor BeneditoNo ratings yet
- A Comprehensive Insight of Current and Future ChalDocument12 pagesA Comprehensive Insight of Current and Future ChalFatima zahra ALTNo ratings yet
- Bodenhausen 2023 Predicting Soil Fungal Communities From CDocument13 pagesBodenhausen 2023 Predicting Soil Fungal Communities From CAlexander RodriguezNo ratings yet
- PhyllosphereDocument38 pagesPhyllosphereRuchi TikarihaNo ratings yet
- Cropping Practices Manipulate Abundance Patterns of Root and Soil MicrobiomeDocument14 pagesCropping Practices Manipulate Abundance Patterns of Root and Soil Microbiomed4rkgr455No ratings yet
- Ecology Letters - 2009 - Fierer - Global Patterns in Belowground CommunitiesDocument12 pagesEcology Letters - 2009 - Fierer - Global Patterns in Belowground CommunitiesFernanda LageNo ratings yet
- Oil PollutionDocument15 pagesOil PollutionDebrup GhoshNo ratings yet
- 2024 - Garzon Lopez Et Al - Remote Sensing Biodiversity Monitoring in Latin AmericaDocument16 pages2024 - Garzon Lopez Et Al - Remote Sensing Biodiversity Monitoring in Latin AmericaAlejandro MirandaNo ratings yet
- The Genetic Architechture of Root and Leaf Microbiota Adaptation in A ThalianaDocument32 pagesThe Genetic Architechture of Root and Leaf Microbiota Adaptation in A ThalianaSergio D. PérezNo ratings yet
- Can Dynamic Network Modelling Be Used ToDocument10 pagesCan Dynamic Network Modelling Be Used ToIlim LivaneliNo ratings yet
- Functional Microbial Ecology Molecular Approaches To MicrobialDocument3 pagesFunctional Microbial Ecology Molecular Approaches To MicrobialLUZ ANDREA GARCIA CAYCEDONo ratings yet
- The Neighbor Avoidance Effect of Microplastics On Ba 2021 Environmental SCDocument9 pagesThe Neighbor Avoidance Effect of Microplastics On Ba 2021 Environmental SCPRAVEEN KUMAR .KNo ratings yet
- Berkelmann Et Al-2020-Environmental Microbiome Compressed-1Document18 pagesBerkelmann Et Al-2020-Environmental Microbiome Compressed-1Anja MeryandiniNo ratings yet
- The Soil Microbiome - From Metagenomics To Metaphenomics: SciencedirectDocument7 pagesThe Soil Microbiome - From Metagenomics To Metaphenomics: SciencedirectMiroz RexNo ratings yet
- 2021 - Rodrigues - Microplastics and Plankton - Knowledge From Laboratory and Field Studies To Distinguish Contamination From PollutionDocument14 pages2021 - Rodrigues - Microplastics and Plankton - Knowledge From Laboratory and Field Studies To Distinguish Contamination From PollutionLim Kok PingNo ratings yet
- The Microbial Connection To Sustainable AgricultureDocument25 pagesThe Microbial Connection To Sustainable AgricultureLAURA VALENTINA RODRIGUEZ ROJASNo ratings yet
- Microorganisms 11 00710Document13 pagesMicroorganisms 11 00710Jull sedanoNo ratings yet
- Soil Food Webs: January 2018Document14 pagesSoil Food Webs: January 2018catnkumar catnkumarNo ratings yet
- Bioaugmentation As A Soil Bioremediation Approach Timothy M VogelDocument6 pagesBioaugmentation As A Soil Bioremediation Approach Timothy M VogelborysNo ratings yet
- Agriculture: Microplastics in Agricultural Systems: Analytical Methodologies and Effects On Soil Quality and Crop YieldDocument29 pagesAgriculture: Microplastics in Agricultural Systems: Analytical Methodologies and Effects On Soil Quality and Crop YieldDaniel ZúñigaNo ratings yet
- New Phytologist - 2019 - Rillig - Microplastic Effects On PlantsDocument5 pagesNew Phytologist - 2019 - Rillig - Microplastic Effects On PlantsSANDRA LUZ MENDOZA LOPEZNo ratings yet
- Mapping The Relative Abundance of Soil Microbiome Biodiversity From eDNA and Remote Sensing - ScienceDirectDocument30 pagesMapping The Relative Abundance of Soil Microbiome Biodiversity From eDNA and Remote Sensing - ScienceDirectHOD PhysicsNo ratings yet
- Phytorem of CD and Microbial Community EffectsDocument10 pagesPhytorem of CD and Microbial Community EffectsValentin dutillieuxNo ratings yet
- Hydroponic Growth Media Substrate A ReviewDocument8 pagesHydroponic Growth Media Substrate A ReviewThan TsafNo ratings yet
- Hill 2000Document12 pagesHill 2000Redd ZhuangNo ratings yet
- Plastic-Degrading Potential Across The Global - 2020Document26 pagesPlastic-Degrading Potential Across The Global - 2020Montagu FryerNo ratings yet
- Saati Santamaría Et Al 2022 Comparative Genomics of The Genus Pseudomonas Reveals Host and Environment SpecificDocument15 pagesSaati Santamaría Et Al 2022 Comparative Genomics of The Genus Pseudomonas Reveals Host and Environment SpecificnfernandezNo ratings yet
- Rapid in Situ Nutrient Element Distribution in Plants and Soils Using Laser Induced Breakdown Spectroscopy (LIBS)Document10 pagesRapid in Situ Nutrient Element Distribution in Plants and Soils Using Laser Induced Breakdown Spectroscopy (LIBS)arturo de miguel granéNo ratings yet
- Omics Breakthroughs For Environmental MicrobiologyDocument16 pagesOmics Breakthroughs For Environmental Microbiologyabcder1234No ratings yet
- Soil Microbial Communities: January 2004Document18 pagesSoil Microbial Communities: January 2004Abdulkadir HassanNo ratings yet
- Bioaugmentation As A Soil Bioremediation Approach Timothy M VogelDocument6 pagesBioaugmentation As A Soil Bioremediation Approach Timothy M VogelnghiasipraNo ratings yet
- Underground Processes 2003Document79 pagesUnderground Processes 2003andresuribeNo ratings yet
- Lebeis 2014Document7 pagesLebeis 2014Francisca Beltrán GuzmánNo ratings yet
- Diversity 14 01122Document15 pagesDiversity 14 01122Stephanie HereiraNo ratings yet
- Culturing The Uncultured Microbial Majority in Activated Sludge: A Critical ReviewDocument26 pagesCulturing The Uncultured Microbial Majority in Activated Sludge: A Critical ReviewmizbaNo ratings yet
- Application of Organic Fertilizer Changes The Rhizosphere Hg7hqjj4Document15 pagesApplication of Organic Fertilizer Changes The Rhizosphere Hg7hqjj4markgomez451No ratings yet
- Mikroplastik Pada Tumbuhan TinggiDocument26 pagesMikroplastik Pada Tumbuhan TinggiarifinmuhammadNo ratings yet
- Biodegradable Mulch Films Significantly Affected Rhizosphere Microbial Communities and Increased Peanut YieldDocument10 pagesBiodegradable Mulch Films Significantly Affected Rhizosphere Microbial Communities and Increased Peanut YieldemerensianaNo ratings yet
- Three Mechanisms of MycoDocument8 pagesThree Mechanisms of MycoAndres LondonoNo ratings yet
- LDPE Microplastics Significantly Alter The Temporal Turnover of Soil Microbial CommunitiesDocument34 pagesLDPE Microplastics Significantly Alter The Temporal Turnover of Soil Microbial CommunitiesYovana Avril Valles AraujoNo ratings yet
- New Phytologist - 2021 - Zhu - Impacts of Global Change On The Phyllosphere MicrobiomeDocument10 pagesNew Phytologist - 2021 - Zhu - Impacts of Global Change On The Phyllosphere MicrobiomeLEIDY TATIANA PEREA PALACIONo ratings yet
- Vigdis Torsvik and Lise Øvreås: Microbial Diversity and Function in Soil: From Genes To EcosystemsDocument6 pagesVigdis Torsvik and Lise Øvreås: Microbial Diversity and Function in Soil: From Genes To EcosystemsSal EmbNo ratings yet
- Molecular Ecology - 2022 - Tedersoo - Best Practices in Metabarcoding of Fungi From Experimental Design To ResultsDocument27 pagesMolecular Ecology - 2022 - Tedersoo - Best Practices in Metabarcoding of Fungi From Experimental Design To Resultsyismar farith mosquera mosqueraNo ratings yet
- Agricultural Soils, Pesticides and Microbial DiversityDocument6 pagesAgricultural Soils, Pesticides and Microbial DiversityAndres DuranNo ratings yet
- 2021 - Paper - Microplastic Residues in Wetland Ecosystems Do They Truly Threaten The Plant-Microbe-Soil SystemDocument12 pages2021 - Paper - Microplastic Residues in Wetland Ecosystems Do They Truly Threaten The Plant-Microbe-Soil SystemMohammad Mehdi NorooziNo ratings yet
- Related Studies - 2Document5 pagesRelated Studies - 2Christian Ivan De LiraNo ratings yet
- Rhizoremediation: A Beneficial Plant-Microbe Interaction: ReviewDocument10 pagesRhizoremediation: A Beneficial Plant-Microbe Interaction: ReviewPritam PatilNo ratings yet
- Otential Health Impacts of Microplastics To Humans and Aquatic EcosystemsDocument14 pagesOtential Health Impacts of Microplastics To Humans and Aquatic EcosystemsPRAVEEN KUMAR .KNo ratings yet
- MetagenomicDocument48 pagesMetagenomicASHIKUZZAMAN ANTORNo ratings yet
- Occurenceandecologicalimpactsof MPinsoilDocument10 pagesOccurenceandecologicalimpactsof MPinsoilSalma ShadNo ratings yet
- Bacteria Estación EspacialDocument17 pagesBacteria Estación EspacialCarlos MendozaNo ratings yet
- Concept Note FinalDocument6 pagesConcept Note Finalanthonyfidelis19No ratings yet
- Comparative Microbiome Analysis of Two Different Long-Term PesticideDocument11 pagesComparative Microbiome Analysis of Two Different Long-Term PesticideCarolina Estuardo NorambuenaNo ratings yet
- Microbial Diversity and Function in Soil-TorvysDocument6 pagesMicrobial Diversity and Function in Soil-TorvysAzucena RosalesNo ratings yet
- Soils as a Key Component of the Critical Zone 6: EcologyFrom EverandSoils as a Key Component of the Critical Zone 6: EcologyPhilippe LemanceauNo ratings yet
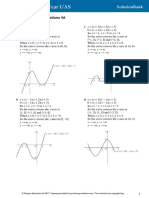
- Continuous Distributions 3A: 1 A Sketching The FunctionDocument8 pagesContinuous Distributions 3A: 1 A Sketching The FunctionAnnaNo ratings yet
- Biomedical Admissions Test 4500/12: Section 2 Scientific Knowledge and ApplicationsDocument20 pagesBiomedical Admissions Test 4500/12: Section 2 Scientific Knowledge and ApplicationsAnnaNo ratings yet
- Oct 2022 iPLS FINALDocument7 pagesOct 2022 iPLS FINALAnnaNo ratings yet
- 1 Key Dates - Updates inDocument6 pages1 Key Dates - Updates inAnnaNo ratings yet
- Interview Podcast - TranscriptDocument5 pagesInterview Podcast - TranscriptAnnaNo ratings yet
- 2022 Examination Dates: Please Note The Following InformationDocument23 pages2022 Examination Dates: Please Note The Following InformationAnnaNo ratings yet
- Sample Oxbridge Interview Questions: GeneralDocument27 pagesSample Oxbridge Interview Questions: GeneralAnna100% (1)
- Overexpression of BHLH Domain of HIF-1 Failed To Inhibit The HIF-1 Transcriptional Activity in HypoxiaDocument11 pagesOverexpression of BHLH Domain of HIF-1 Failed To Inhibit The HIF-1 Transcriptional Activity in HypoxiaAnnaNo ratings yet
- Why The Zebra Got Its Stripes: To Deter Flies From Landing On ItDocument4 pagesWhy The Zebra Got Its Stripes: To Deter Flies From Landing On ItAnnaNo ratings yet
- Why Does A Zebra Have Stripes?: (Maybe This Is The Wrong Question)Document7 pagesWhy Does A Zebra Have Stripes?: (Maybe This Is The Wrong Question)AnnaNo ratings yet
- Knockdown of Long Non-Coding Rna Hotair Reverses Cisplatin Resistance of Ovarian Cancer Cells Through Inhibiting Mir-138-5P-Regulated Ezh2 and Sirt1Document10 pagesKnockdown of Long Non-Coding Rna Hotair Reverses Cisplatin Resistance of Ovarian Cancer Cells Through Inhibiting Mir-138-5P-Regulated Ezh2 and Sirt1AnnaNo ratings yet
- AAMC Standardized Immunization Form: MMR - 2 Doses of MMRDocument4 pagesAAMC Standardized Immunization Form: MMR - 2 Doses of MMRAnnaNo ratings yet
- Biomedical Admissions Test (Bmat) Registration Form OCTOBER 2019Document5 pagesBiomedical Admissions Test (Bmat) Registration Form OCTOBER 2019AnnaNo ratings yet
- Instructions For Completing The 2020-21 GRE Fee Reduction Request FormDocument3 pagesInstructions For Completing The 2020-21 GRE Fee Reduction Request FormAnnaNo ratings yet
- 2020-21 GRE Subject Tests Registration FormDocument6 pages2020-21 GRE Subject Tests Registration FormAnnaNo ratings yet
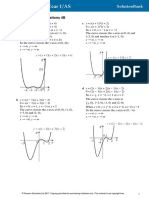
- Graphs and Transformations 4A: 1 A y (X C y (X + 1) (X + 2) (X + 3)Document11 pagesGraphs and Transformations 4A: 1 A y (X C y (X + 1) (X + 2) (X + 3)AnnaNo ratings yet
- Alevelsb p1 Ex3bDocument4 pagesAlevelsb p1 Ex3bAnnaNo ratings yet
- Equations and Inequalities 3CDocument2 pagesEquations and Inequalities 3CAnnaNo ratings yet
- Graphs and Transformations 4B: 1 A y (X + 1) (X + 2) (X + 3) (X + 4) C y X (X + 1)Document4 pagesGraphs and Transformations 4B: 1 A y (X + 1) (X + 2) (X + 3) (X + 4) C y X (X + 1)AnnaNo ratings yet
- Algebraic Expressions 1C: y 3xy (3y + 4x)Document2 pagesAlgebraic Expressions 1C: y 3xy (3y + 4x)AnnaNo ratings yet
- Equations and Inequalities 3D: X X X X X X XDocument3 pagesEquations and Inequalities 3D: X X X X X X XAnnaNo ratings yet
- Equations and Inequalities 3A: 1 A Multiply 2x D So Solution Is e Multiply 3xDocument2 pagesEquations and Inequalities 3A: 1 A Multiply 2x D So Solution Is e Multiply 3xAnnaNo ratings yet
- Alevelsb p1 Ex2cDocument1 pageAlevelsb p1 Ex2cAnnaNo ratings yet
- CH 9Document8 pagesCH 9Maulik ChaudharyNo ratings yet
- International Journal of Nursing Studies: ArticleinfoDocument10 pagesInternational Journal of Nursing Studies: ArticleinfoJihanMukhtarNo ratings yet
- Final Syllabus B Planning 2017 SPA DelhiDocument44 pagesFinal Syllabus B Planning 2017 SPA DelhiGATE AR97No ratings yet
- Kristina Wirtz Ritual, Discourse, and Community in Cuban Santeria Speaking A Sacred WorldDocument281 pagesKristina Wirtz Ritual, Discourse, and Community in Cuban Santeria Speaking A Sacred WorldDiego Sebastián Rojas Toro100% (1)
- Set P4 TGDocument228 pagesSet P4 TGYumvuhore boscoNo ratings yet
- Cognitive PsychologyDocument10 pagesCognitive Psychologydominscience1No ratings yet
- AuthorDocument25 pagesAuthorJae anne UdtohanNo ratings yet
- B2B Buying Behavior.01Document16 pagesB2B Buying Behavior.01kumar shashank keshri100% (1)
- SIMCARDDocument16 pagesSIMCARDJocelle BilagNo ratings yet
- Ex HypothesistestDocument22 pagesEx HypothesistestYana CovarNo ratings yet
- Impact of Inter Vehicle Communication (IVC) On Network and Road Traffic Simulation Using Bidirectional Coupled Network Through One Hop Broadcasting TechniqueDocument6 pagesImpact of Inter Vehicle Communication (IVC) On Network and Road Traffic Simulation Using Bidirectional Coupled Network Through One Hop Broadcasting TechniqueseventhsensegroupNo ratings yet
- Evaluation of Training & RetrainingDocument3 pagesEvaluation of Training & RetrainingNischal ChhabriaNo ratings yet
- Factors Affecting Stress Among Faculty Members ofDocument15 pagesFactors Affecting Stress Among Faculty Members ofMichevelli RiveraNo ratings yet
- Sample Thesis Chapter 1 Statement of The ProblemDocument4 pagesSample Thesis Chapter 1 Statement of The ProblemOrderAPaperUK100% (2)
- 1.6.10.3-ASTM-C823-Standard Practice For Examinations & SamplingDocument7 pages1.6.10.3-ASTM-C823-Standard Practice For Examinations & SamplingmanikNo ratings yet
- Minitab Problem 6.1Document2 pagesMinitab Problem 6.1Varsha MalviyaNo ratings yet
- E. Bachelor Degree Project I Supervisor Evaluation FormDocument1 pageE. Bachelor Degree Project I Supervisor Evaluation FormDex JHNo ratings yet
- Week 5Document10 pagesWeek 5Coco LlameraNo ratings yet
- Use of Calculators in ExaminationsDocument3 pagesUse of Calculators in ExaminationsrohanNo ratings yet
- Global Strategic ManagementDocument4 pagesGlobal Strategic ManagementDrYogesh MehtaNo ratings yet
- Sampling Theory - NotesDocument43 pagesSampling Theory - NotesAnand Huralikoppi100% (2)
- Geometric Correction For Landsat ImaginaryDocument23 pagesGeometric Correction For Landsat ImaginaryIvan BarriaNo ratings yet
- Autoencoder Asset Pricing ModelsDocument22 pagesAutoencoder Asset Pricing ModelsEdson KitaniNo ratings yet
- Concept of GuidanceDocument16 pagesConcept of GuidanceAkiraNo ratings yet
- Keywords: Service Quality, Return Intention, Peak Natural EnchantmentDocument16 pagesKeywords: Service Quality, Return Intention, Peak Natural EnchantmentWinson LieNo ratings yet
- Bloom's TaxonomyDocument15 pagesBloom's Taxonomyrmit_998186100% (2)
- Notice: Draft Guidance For Industry: Validation of Growth-Based Rapid Microbiological Methods For Sterility Testing of Cellular and Gene Therapy Products AvailabilityDocument2 pagesNotice: Draft Guidance For Industry: Validation of Growth-Based Rapid Microbiological Methods For Sterility Testing of Cellular and Gene Therapy Products AvailabilityJustia.com100% (4)
- Work SamplingDocument25 pagesWork Samplingkunwaraditya100% (1)