Download as pdf or txt
You might also like
- Cancer Biology 2023Document82 pagesCancer Biology 2023STACEY SALVILLANo ratings yet
- Tumor MarkersDocument145 pagesTumor MarkersAnsh Dutta50% (2)
- Bishwarup SarkarDocument18 pagesBishwarup SarkarAnyone Can CookNo ratings yet
- Role of Hypoxia in Cancer Therapy by Regulating The Tumor MicroenvironmentDocument15 pagesRole of Hypoxia in Cancer Therapy by Regulating The Tumor MicroenvironmentShuang LiNo ratings yet
- Research Paper 142CDocument16 pagesResearch Paper 142CseydNo ratings yet
- Cells 10 03017 v2Document38 pagesCells 10 03017 v2ANDREA BERENICE ARMENTA RAZONo ratings yet
- Protac 2Document14 pagesProtac 2muruganvvkNo ratings yet
- Targeting Hypoxia in The Tumor Microenvironment ADocument16 pagesTargeting Hypoxia in The Tumor Microenvironment AViviana OrellanaNo ratings yet
- s12916 022 02444 8Document14 pagess12916 022 02444 8Dr Meenakshi ParwaniNo ratings yet
- Journal of Pineal Research - 2007 - Dai - Melatonin Modulates The Expression of VEGF and HIF 1 Induced by CoCl2 inDocument6 pagesJournal of Pineal Research - 2007 - Dai - Melatonin Modulates The Expression of VEGF and HIF 1 Induced by CoCl2 inADRIANANo ratings yet
- Bremer 2017Document11 pagesBremer 2017ziNo ratings yet
- 2022 BMC Cancer 22 1211Document17 pages2022 BMC Cancer 22 1211mbrylinskiNo ratings yet
- Oxygen and Viruses: A Breathing Story: ReviewDocument4 pagesOxygen and Viruses: A Breathing Story: ReviewVedanthNo ratings yet
- HDAC3 Inhibition Up Regulates PD-L1 Expression in B-Cell Lymphomas and Augments The Efficacy of Anti-PD-L1 TherapyDocument28 pagesHDAC3 Inhibition Up Regulates PD-L1 Expression in B-Cell Lymphomas and Augments The Efficacy of Anti-PD-L1 Therapyshidis1028No ratings yet
- Relevant Pharmacological Interactions Between Alkylating Agents and Antiepileptic Drugs: Preclinical and Clinical DataDocument13 pagesRelevant Pharmacological Interactions Between Alkylating Agents and Antiepileptic Drugs: Preclinical and Clinical DataFederico CucchiaraNo ratings yet
- Research Article Pharmacotherapeutics and Molecular Mechanism of Phytochemicals in Alleviating Hormone-Responsive Breast CancerDocument15 pagesResearch Article Pharmacotherapeutics and Molecular Mechanism of Phytochemicals in Alleviating Hormone-Responsive Breast CancerdinaNo ratings yet
- Akinyele Et Al 2023 Investigation of Methyl 5 Pentan 3 Yloxy 7 Oxabicyclo 4 1 0 Hept 3 Ene 3 CarboxyhydrazideDocument12 pagesAkinyele Et Al 2023 Investigation of Methyl 5 Pentan 3 Yloxy 7 Oxabicyclo 4 1 0 Hept 3 Ene 3 CarboxyhydrazideOmolara AdeboyeNo ratings yet
- Agriculture and Natural ResourcesDocument10 pagesAgriculture and Natural ResourcesSmonagenes UbNo ratings yet
- Biochemical and Biophysical Research CommunicationsDocument7 pagesBiochemical and Biophysical Research Communicationslecture fkNo ratings yet
- Zhou Et Al 2023Document10 pagesZhou Et Al 2023Caique SilvaNo ratings yet
- 2021 Article 1063Document15 pages2021 Article 1063Raúl LópezNo ratings yet
- Addition of A Cholesterol Group To An HIV-1 Peptide Fusion Inhibitor Dramatically Increases Its Antiviral PotencyDocument6 pagesAddition of A Cholesterol Group To An HIV-1 Peptide Fusion Inhibitor Dramatically Increases Its Antiviral PotencyGmo AnraNo ratings yet
- Quenching of Quorum Sensing in Multi-Drug Resistant Pseudomonas Aeruginosa Insights On Halo-Bacterial Metabolites and Gamma Irradiation As Channels InhibitorsDocument15 pagesQuenching of Quorum Sensing in Multi-Drug Resistant Pseudomonas Aeruginosa Insights On Halo-Bacterial Metabolites and Gamma Irradiation As Channels Inhibitorsd4rkgr455No ratings yet
- Vooijs2008 - Metastasis - Fig3Document9 pagesVooijs2008 - Metastasis - Fig3Nisa TanNo ratings yet
- tmp9CE5 TMPDocument8 pagestmp9CE5 TMPFrontiersNo ratings yet
- Zhou Et Al 2024 The Small Molecule CBR 5884 Inhibits The Candida Albicans Phosphatidylserine SynthaseDocument23 pagesZhou Et Al 2024 The Small Molecule CBR 5884 Inhibits The Candida Albicans Phosphatidylserine SynthaseMichael UNo ratings yet
- Long-Term Prophylaxis With Androgens in The Management of Hereditary Angioedema (HAE) in Emerging CountriesDocument8 pagesLong-Term Prophylaxis With Androgens in The Management of Hereditary Angioedema (HAE) in Emerging CountriesAthar 1101No ratings yet
- A Randomized Trial Evaluating The Association Between Related Gene Polymorphism and Nausea and Vomiting Induced by Cisplatin Multi-Day ChemotherapyDocument10 pagesA Randomized Trial Evaluating The Association Between Related Gene Polymorphism and Nausea and Vomiting Induced by Cisplatin Multi-Day ChemotherapyGABRIELANo ratings yet
- Yjbm 80 2 51Document10 pagesYjbm 80 2 51dana ursuNo ratings yet
- The FEBS Journal - 2020 - Vanderhaeghen - Hypoxia Inducible Factors in Metabolic Reprogramming During SepsisDocument18 pagesThe FEBS Journal - 2020 - Vanderhaeghen - Hypoxia Inducible Factors in Metabolic Reprogramming During SepsisADRIANANo ratings yet
- Hif-1 Inibidores NaturaisDocument18 pagesHif-1 Inibidores NaturaisJoão FrancoNo ratings yet
- Baw 159Document10 pagesBaw 159Karim KACEMNo ratings yet
- 2020 Article 1654Document13 pages2020 Article 1654alyaNo ratings yet
- Therapeutic Application of Monoclonal Antibodies: A Review: September 2020Document17 pagesTherapeutic Application of Monoclonal Antibodies: A Review: September 2020segun shonaNo ratings yet
- Poly ε-Caprolactone Nanoparticles for Sustained Intra-Articular Immune Modulation in Adjuvant-Induced Arthritis Rodent ModelDocument22 pagesPoly ε-Caprolactone Nanoparticles for Sustained Intra-Articular Immune Modulation in Adjuvant-Induced Arthritis Rodent ModelAman DubeyNo ratings yet
- 2022 The Effects of Hyperbaric Oxygen Therapy (HBOT) On Coronavirus Disease-2019 (COVID-19) - A Systematic ReviewDocument12 pages2022 The Effects of Hyperbaric Oxygen Therapy (HBOT) On Coronavirus Disease-2019 (COVID-19) - A Systematic ReviewabnfojoNo ratings yet
- 1 s2.0 S1476927118301087 MainDocument12 pages1 s2.0 S1476927118301087 MainArumNo ratings yet
- The Role of Hypoxia-Inducible Factor-1alpha and Its Signaling in MelanomaDocument9 pagesThe Role of Hypoxia-Inducible Factor-1alpha and Its Signaling in Melanomamohammad malekanNo ratings yet
- Bepridil Is Potent Against SARS-CoV-2 in VitroDocument28 pagesBepridil Is Potent Against SARS-CoV-2 in VitrosovalaxNo ratings yet
- RO5 J Ijpharm 2018 07 046Document60 pagesRO5 J Ijpharm 2018 07 046Dina AzkiyahNo ratings yet
- 403535v1 FullDocument25 pages403535v1 FullFahmi FadilahNo ratings yet
- Multi-Targeted Therapy Resistance Via Drug - Induced Secretome FucosylationDocument42 pagesMulti-Targeted Therapy Resistance Via Drug - Induced Secretome Fucosylationaditya subchanNo ratings yet
- Mitochondrial Adaptation in Cancer Drug ResistanceDocument42 pagesMitochondrial Adaptation in Cancer Drug ResistanceporelsudiptaNo ratings yet
- Assays For InteractionsDocument16 pagesAssays For InteractionsRam KNo ratings yet
- Tnfpred: Identifying Tumor Necrosis Factors Using Hybrid Features Based On Word EmbeddingsDocument11 pagesTnfpred: Identifying Tumor Necrosis Factors Using Hybrid Features Based On Word EmbeddingsNguyen Trinh Trung DuongNo ratings yet
- s12888 023 05324 4Document14 pagess12888 023 05324 4Ronaldo OliveiraNo ratings yet
- 2011 04 Clin Cancer Res - AfzalDocument9 pages2011 04 Clin Cancer Res - AfzalEspen Jimenez SolemNo ratings yet
- Virtual Screening of Phytochemical Compounds As Potential Inhibitors Against HIV-1 Resistance To Protease InhibitorDocument6 pagesVirtual Screening of Phytochemical Compounds As Potential Inhibitors Against HIV-1 Resistance To Protease InhibitorZoya AmjadNo ratings yet
- Fabrication of Hesperidin Nanoparticles Loaded by Poly Lactic Co-Glycolic Acid For Improved Therapeutic Efficiency and CytotoxicityDocument18 pagesFabrication of Hesperidin Nanoparticles Loaded by Poly Lactic Co-Glycolic Acid For Improved Therapeutic Efficiency and CytotoxicityHoàng Sơn Nguyễn LêNo ratings yet
- Anti-Drug AntibodiesDocument8 pagesAnti-Drug AntibodiesDrugs & Therapy StudiesNo ratings yet
- Pharmaceutical Sciences: Targeting Hif-1 Pathway: A Therapeutic Approach To Kill Cancer CellsDocument6 pagesPharmaceutical Sciences: Targeting Hif-1 Pathway: A Therapeutic Approach To Kill Cancer CellsBaru Chandrasekhar RaoNo ratings yet
- New Mechanisms of Radioiodide Uptake Revealed Via A Novel High Throughput Drug Screening Approach in Thyroid CancerDocument26 pagesNew Mechanisms of Radioiodide Uptake Revealed Via A Novel High Throughput Drug Screening Approach in Thyroid CancerWalid SasiNo ratings yet
- Viruses 15 00804 With Cover 1Document25 pagesViruses 15 00804 With Cover 1acheikhrouhouNo ratings yet
- s13063 021 05224 6Document11 pagess13063 021 05224 6Nejc KovačNo ratings yet
- The Antityrosinase and Antioxidant Activities of Flavonoids Dominated by The Number and Location of Phenolic Hydroxyl GroupsDocument12 pagesThe Antityrosinase and Antioxidant Activities of Flavonoids Dominated by The Number and Location of Phenolic Hydroxyl GroupsRico RamadhanNo ratings yet
- Editorials: Hyperbaric Oxygen For Invasive Fungal InfectionsDocument2 pagesEditorials: Hyperbaric Oxygen For Invasive Fungal InfectionsFahmee MujaheedNo ratings yet
- CLEOPATRA Biomarker Analysis - ProtocolDocument117 pagesCLEOPATRA Biomarker Analysis - ProtocolweilinmdNo ratings yet
- The Application of Histone Deacetylases Inhibitors in GlioblastomaDocument18 pagesThe Application of Histone Deacetylases Inhibitors in GlioblastomaruthameliapNo ratings yet
- Efficacy and Safety Profile of Ivosidenib in The Management of Patients With Acute Myeloid Leukemia AML An Update On The Emerging EvidenceDocument15 pagesEfficacy and Safety Profile of Ivosidenib in The Management of Patients With Acute Myeloid Leukemia AML An Update On The Emerging EvidenceBismillah BerkahNo ratings yet
- Antioxidants Stimulate BACH1-dependent Tumor AngiogenesisDocument34 pagesAntioxidants Stimulate BACH1-dependent Tumor AngiogenesisA ANo ratings yet
- Next Generation Kinase Inhibitors: Moving Beyond the ATP Binding/Catalytic SitesFrom EverandNext Generation Kinase Inhibitors: Moving Beyond the ATP Binding/Catalytic SitesNo ratings yet
- Pharmaceutical Biotechnology: Fundamentals and ApplicationsFrom EverandPharmaceutical Biotechnology: Fundamentals and ApplicationsRating: 5 out of 5 stars5/5 (1)
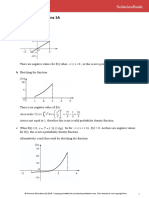
- Continuous Distributions 3A: 1 A Sketching The FunctionDocument8 pagesContinuous Distributions 3A: 1 A Sketching The FunctionAnnaNo ratings yet
- Biomedical Admissions Test 4500/12: Section 2 Scientific Knowledge and ApplicationsDocument20 pagesBiomedical Admissions Test 4500/12: Section 2 Scientific Knowledge and ApplicationsAnnaNo ratings yet
- 1 Key Dates - Updates inDocument6 pages1 Key Dates - Updates inAnnaNo ratings yet
- Microbial Communities From Arid Environments On A Global Scale. A Systematic ReviewDocument12 pagesMicrobial Communities From Arid Environments On A Global Scale. A Systematic ReviewAnnaNo ratings yet
- 2022 Examination Dates: Please Note The Following InformationDocument23 pages2022 Examination Dates: Please Note The Following InformationAnnaNo ratings yet
- Sample Oxbridge Interview Questions: GeneralDocument27 pagesSample Oxbridge Interview Questions: GeneralAnna100% (1)
- Oct 2022 iPLS FINALDocument7 pagesOct 2022 iPLS FINALAnnaNo ratings yet
- Knockdown of Long Non-Coding Rna Hotair Reverses Cisplatin Resistance of Ovarian Cancer Cells Through Inhibiting Mir-138-5P-Regulated Ezh2 and Sirt1Document10 pagesKnockdown of Long Non-Coding Rna Hotair Reverses Cisplatin Resistance of Ovarian Cancer Cells Through Inhibiting Mir-138-5P-Regulated Ezh2 and Sirt1AnnaNo ratings yet
- Interview Podcast - TranscriptDocument5 pagesInterview Podcast - TranscriptAnnaNo ratings yet
- AAMC Standardized Immunization Form: MMR - 2 Doses of MMRDocument4 pagesAAMC Standardized Immunization Form: MMR - 2 Doses of MMRAnnaNo ratings yet
- Why Does A Zebra Have Stripes?: (Maybe This Is The Wrong Question)Document7 pagesWhy Does A Zebra Have Stripes?: (Maybe This Is The Wrong Question)AnnaNo ratings yet
- Why The Zebra Got Its Stripes: To Deter Flies From Landing On ItDocument4 pagesWhy The Zebra Got Its Stripes: To Deter Flies From Landing On ItAnnaNo ratings yet
- 2020-21 GRE Subject Tests Registration FormDocument6 pages2020-21 GRE Subject Tests Registration FormAnnaNo ratings yet
- Algebraic Expressions 1C: y 3xy (3y + 4x)Document2 pagesAlgebraic Expressions 1C: y 3xy (3y + 4x)AnnaNo ratings yet
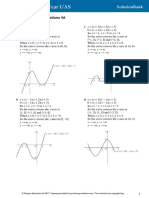
- Graphs and Transformations 4B: 1 A y (X + 1) (X + 2) (X + 3) (X + 4) C y X (X + 1)Document4 pagesGraphs and Transformations 4B: 1 A y (X + 1) (X + 2) (X + 3) (X + 4) C y X (X + 1)AnnaNo ratings yet
- Instructions For Completing The 2020-21 GRE Fee Reduction Request FormDocument3 pagesInstructions For Completing The 2020-21 GRE Fee Reduction Request FormAnnaNo ratings yet
- Biomedical Admissions Test (Bmat) Registration Form OCTOBER 2019Document5 pagesBiomedical Admissions Test (Bmat) Registration Form OCTOBER 2019AnnaNo ratings yet
- Alevelsb p1 Ex3bDocument4 pagesAlevelsb p1 Ex3bAnnaNo ratings yet
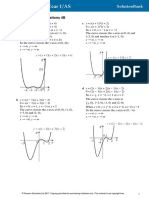
- Graphs and Transformations 4A: 1 A y (X C y (X + 1) (X + 2) (X + 3)Document11 pagesGraphs and Transformations 4A: 1 A y (X C y (X + 1) (X + 2) (X + 3)AnnaNo ratings yet
- Equations and Inequalities 3CDocument2 pagesEquations and Inequalities 3CAnnaNo ratings yet
- Equations and Inequalities 3D: X X X X X X XDocument3 pagesEquations and Inequalities 3D: X X X X X X XAnnaNo ratings yet
- Alevelsb p1 Ex2cDocument1 pageAlevelsb p1 Ex2cAnnaNo ratings yet
- Equations and Inequalities 3A: 1 A Multiply 2x D So Solution Is e Multiply 3xDocument2 pagesEquations and Inequalities 3A: 1 A Multiply 2x D So Solution Is e Multiply 3xAnnaNo ratings yet
- Naringenin A Potential FlavonoidDocument19 pagesNaringenin A Potential FlavonoidLuciano SartiniNo ratings yet
- Neutrophils in Pancreatic Cancer ProgressionDocument103 pagesNeutrophils in Pancreatic Cancer ProgressionShahid ShaikhNo ratings yet
- Dr. Diah Rumekti hadiatiAPCGS JogjaDocument25 pagesDr. Diah Rumekti hadiatiAPCGS JogjacirererereNo ratings yet
- Dumas Et Al., 2021Document16 pagesDumas Et Al., 2021Miguel Ángel Olivas AguirreNo ratings yet
- Umftgm 2017 MarissiensisDocument148 pagesUmftgm 2017 MarissiensisAdriana VillarrealNo ratings yet
- Understanding and Leveraging Phenotypic PlasticityDocument11 pagesUnderstanding and Leveraging Phenotypic Plasticitysneha.921530sharmaNo ratings yet
- Literature Review On Vital SignsDocument6 pagesLiterature Review On Vital Signsafdtwbhyk100% (1)
- Protective Effects of Blue Light-Blocking Shades On Phototoxicity in Human Ocular Surface CellsDocument6 pagesProtective Effects of Blue Light-Blocking Shades On Phototoxicity in Human Ocular Surface CellsHans Steven KurniawanNo ratings yet
- June Web FriendlyDocument76 pagesJune Web FriendlyИван ХрищенюкNo ratings yet
- Kuriakose Contemporary Oral Oncology Biology, Epidemiology, EtiologyDocument546 pagesKuriakose Contemporary Oral Oncology Biology, Epidemiology, Etiologyخالد الجدياويNo ratings yet
- Web of Science (5.13.1) - Export Transfer ServiceDocument17 pagesWeb of Science (5.13.1) - Export Transfer ServicealbertosNo ratings yet
- Circulating Tumor Cells in Breast Cancer Metastatic DiseaseDocument177 pagesCirculating Tumor Cells in Breast Cancer Metastatic DiseaseAd AdrianaNo ratings yet
- Scientific Programme-TCR2020 PDFDocument13 pagesScientific Programme-TCR2020 PDFLipsa SahooNo ratings yet
- 1 s2.0 S235230421930100X Main PDFDocument14 pages1 s2.0 S235230421930100X Main PDFLorena RamosNo ratings yet
- Jurnal KolangiokarsinomaDocument15 pagesJurnal KolangiokarsinomaAnonymous cwlpSlReUYNo ratings yet
- Defining Epidermal Basal Cell States During Skin Homeostasis and Wound Healing Using Single-Cell TranscriptomicsDocument23 pagesDefining Epidermal Basal Cell States During Skin Homeostasis and Wound Healing Using Single-Cell TranscriptomicsSonia CaruntuNo ratings yet
- Modeling Heterogeneity 2022Document12 pagesModeling Heterogeneity 2022Shuang LiNo ratings yet
- Materials Today Bio: Jiahao Wang, Peiyi Zhao, Zhirong Chen, Hui Wang, Yajia Wang, Quankui LinDocument15 pagesMaterials Today Bio: Jiahao Wang, Peiyi Zhao, Zhirong Chen, Hui Wang, Yajia Wang, Quankui Lin马三强No ratings yet
- Ijmm 31 6 1375 PDFDocument6 pagesIjmm 31 6 1375 PDFAlex RogobeteNo ratings yet
- Research Paper EndingDocument7 pagesResearch Paper Endingafeaaawci100% (1)
- Frontal Fibrosing Alopecia An Update On The Hy - 2019 - International Journal oDocument8 pagesFrontal Fibrosing Alopecia An Update On The Hy - 2019 - International Journal oFlamina DerniNo ratings yet
- Articulo 2Document17 pagesArticulo 2JL MaravillasNo ratings yet
- Magee2017 Article DiabeticNephropathyATangledWebDocument14 pagesMagee2017 Article DiabeticNephropathyATangledWebYulian 53No ratings yet
- Pharmacological ResearchDocument12 pagesPharmacological ResearchJUAN DIEGO TRUJILLO ROJASNo ratings yet
- Jurnal NeoplasmaDocument10 pagesJurnal NeoplasmaFadhilah Asyifa DewantiNo ratings yet
- Caramori 2019Document16 pagesCaramori 2019Dusan MilovanovicNo ratings yet
- Research Profiles For KUMCDocument96 pagesResearch Profiles For KUMC김수함No ratings yet
- Immunohistochemical Expression of CD44 and CD56 in Malignant Salivary Gland TumorsDocument12 pagesImmunohistochemical Expression of CD44 and CD56 in Malignant Salivary Gland TumorsIJAR JOURNALNo ratings yet