Download as pdf or txt
You might also like
- Impact 2 Student S BookDocument165 pagesImpact 2 Student S Bookmaria83% (6)
- Paediatric Chest RadiologyDocument120 pagesPaediatric Chest Radiologysunielgoel100% (6)
- Food Microbiology: Dane A. Jensen, Loretta M. Friedrich, Linda J. Harris, Michelle D. Danyluk, Donald W. SchaffnerDocument6 pagesFood Microbiology: Dane A. Jensen, Loretta M. Friedrich, Linda J. Harris, Michelle D. Danyluk, Donald W. SchaffnerJaime Lagos BorquezNo ratings yet
- Transfer of Escherichia Coli O157:H7 From Soil, Water, and Manure Contaminated With Low Numbers of The Pathogen To Lettuce PlantsDocument5 pagesTransfer of Escherichia Coli O157:H7 From Soil, Water, and Manure Contaminated With Low Numbers of The Pathogen To Lettuce PlantsDini Gustiari TondangNo ratings yet
- Microorganisms 11 01801 v2Document20 pagesMicroorganisms 11 01801 v2hmgaparaisoNo ratings yet
- The Survival of Escherichia Coli O157:H7 in The Presence of Penicillium Expansum and Glomerella Cingulata in Wounds On Apple SurfacesDocument7 pagesThe Survival of Escherichia Coli O157:H7 in The Presence of Penicillium Expansum and Glomerella Cingulata in Wounds On Apple SurfacesMarwa aljarahNo ratings yet
- Food Microbiology: Short CommunicationDocument4 pagesFood Microbiology: Short CommunicationAustin KlasnerNo ratings yet
- Research Communications: Influence of Different Strains of Induction of Hairy Roots and Artemisinin Production inDocument5 pagesResearch Communications: Influence of Different Strains of Induction of Hairy Roots and Artemisinin Production inDương ThùyNo ratings yet
- Lytic Bacteriophages Reduce Escherichia Coli O157:H7 On Fresh Cut Lettuce Introduced Through Cross-ContaminationDocument8 pagesLytic Bacteriophages Reduce Escherichia Coli O157:H7 On Fresh Cut Lettuce Introduced Through Cross-ContaminationTany@No ratings yet
- CONCLUSION-Lab Report MicroDocument2 pagesCONCLUSION-Lab Report Microfdhln sakinahNo ratings yet
- Chen 2019Document11 pagesChen 2019Kornkamon SriprasertNo ratings yet
- 2014.1.5.systemic Evaluation of Antibacterial Activity ofDocument7 pages2014.1.5.systemic Evaluation of Antibacterial Activity offeNo ratings yet
- Capsicum Annuum: Antibacterial Activities of Endophytic Fungi From L. (Siling Labuyo) Leaves and FruitsDocument10 pagesCapsicum Annuum: Antibacterial Activities of Endophytic Fungi From L. (Siling Labuyo) Leaves and FruitsMeynard Angelo M. JavierNo ratings yet
- Food Con 29 (2013) 318-327Document10 pagesFood Con 29 (2013) 318-327Antonia GounadakiNo ratings yet
- Semisi PPathol 89Document7 pagesSemisi PPathol 89melvinprasad1984No ratings yet
- Dual Role of Endophytic Entomopathogenic Fungi Induce Plant Growth andDocument29 pagesDual Role of Endophytic Entomopathogenic Fungi Induce Plant Growth andEsther AguileraNo ratings yet
- Yun2015 PestalotiopsisDocument6 pagesYun2015 PestalotiopsisHéctor Gómez YáñezNo ratings yet
- Artículo Microorganismos PatogenosDocument12 pagesArtículo Microorganismos PatogenosmanuelaortizbarbosaNo ratings yet
- Plant ArticleDocument5 pagesPlant ArticlevijayselvendranNo ratings yet
- 2019 - Molecular Characteristics and Genotyping Diversity of EHEC O157H7 Isoates in Gauteng Resion South AfricaDocument8 pages2019 - Molecular Characteristics and Genotyping Diversity of EHEC O157H7 Isoates in Gauteng Resion South AfricamotibaNo ratings yet
- 101 Propagation of Juglans Cinerea L Butternut PDFDocument62 pages101 Propagation of Juglans Cinerea L Butternut PDFtalitha nabilla wNo ratings yet
- Iyrc2021 38 FinalDocument9 pagesIyrc2021 38 FinalNatalia ArevaloNo ratings yet
- Plant Growth and Development (Ncert Solution)Document10 pagesPlant Growth and Development (Ncert Solution)Ayush DuttaNo ratings yet
- Bacterias en LechugasDocument9 pagesBacterias en LechugasLuisa PadillaNo ratings yet
- Identification and Characterization of PDocument14 pagesIdentification and Characterization of PlynneunlocksNo ratings yet
- Endophytic Bacteria ThesisDocument4 pagesEndophytic Bacteria Thesisbcrqhr1n100% (2)
- Antibacterial Activity of Garlic Extract Against Human Pathogenic Bacteria 2329 6887 1000253 PDFDocument5 pagesAntibacterial Activity of Garlic Extract Against Human Pathogenic Bacteria 2329 6887 1000253 PDFCiprian CorneaNo ratings yet
- J Ijfoodmicro 2005 02 012Document10 pagesJ Ijfoodmicro 2005 02 012kom dehNo ratings yet
- 93572-Article Text-122181-1-10-20170607Document5 pages93572-Article Text-122181-1-10-20170607Jaru JackyNo ratings yet
- Bioactive Endophytes Warrant Intensified Exploration and ConservationDocument4 pagesBioactive Endophytes Warrant Intensified Exploration and ConservationNurul Robiatul AdawiyahNo ratings yet
- LatayadaFelmer Published PaperDocument5 pagesLatayadaFelmer Published PaperalmairahNo ratings yet
- JWorldPoultRes9107 142019Document9 pagesJWorldPoultRes9107 14201990210No ratings yet
- Biochemical Study For The E Ect of Henna (Lawsonia Inermis) On Escherichia ColiDocument9 pagesBiochemical Study For The E Ect of Henna (Lawsonia Inermis) On Escherichia ColiTuấn Nguyen AnhNo ratings yet
- Isolation, Diversity, and Growth-Promoting Activities of Endophytic Bacteria From Tea Cultivars of Zijuan and Yunkang-10Document11 pagesIsolation, Diversity, and Growth-Promoting Activities of Endophytic Bacteria From Tea Cultivars of Zijuan and Yunkang-10maczweiNo ratings yet
- Physiology of Domesticated Alliums Onions, Garlic, Leek, and Minor CropsDocument7 pagesPhysiology of Domesticated Alliums Onions, Garlic, Leek, and Minor CropsMaria FonsecaNo ratings yet
- Pseudomonas - Cichorii - As - The - Causal - Agent20160830 29498 Qupkhk With Cover Page v2Document17 pagesPseudomonas - Cichorii - As - The - Causal - Agent20160830 29498 Qupkhk With Cover Page v2MarianaNo ratings yet
- Effect of Seed Treatment On Seedling Health of Chili: M. Z. Alam, I. Hamim, M. A. Ali, and M. AshrafuzzamanDocument5 pagesEffect of Seed Treatment On Seedling Health of Chili: M. Z. Alam, I. Hamim, M. A. Ali, and M. Ashrafuzzamanyasir majeedNo ratings yet
- Integration of Biological and Chemical Controls For Rhizoctonia Aerial Blight and Root Rot of RosemaryDocument4 pagesIntegration of Biological and Chemical Controls For Rhizoctonia Aerial Blight and Root Rot of RosemaryDan Kielvin LascanoNo ratings yet
- Article WJPR 1551403980Document10 pagesArticle WJPR 1551403980Dharmvir ChouhanNo ratings yet
- crmb457 4611Document6 pagescrmb457 4611as1as2as3No ratings yet
- Appl. Environ. Microbiol.-2005-Compant-1685-93Document9 pagesAppl. Environ. Microbiol.-2005-Compant-1685-93Lucas Antonio GallaratoNo ratings yet
- Antimicrobial Activities of An Oxygenated Cyclohexanone Derivative Isolated FromDocument36 pagesAntimicrobial Activities of An Oxygenated Cyclohexanone Derivative Isolated FromUtari ZulfianiNo ratings yet
- 450d PDFDocument6 pages450d PDFKyle BarzaNo ratings yet
- tmp7CEC TMPDocument8 pagestmp7CEC TMPFrontiersNo ratings yet
- Studies On Biodiversity of Phylloplane and Endophytic Fungi From Mature Leaves of Medicinal Plant, Carica Papaya L. With Antimicrobial PotentialDocument6 pagesStudies On Biodiversity of Phylloplane and Endophytic Fungi From Mature Leaves of Medicinal Plant, Carica Papaya L. With Antimicrobial PotentialRanadive AnanthNo ratings yet
- Tomato Tissue Culture ThesisDocument6 pagesTomato Tissue Culture ThesisOnlinePaperWritingServicesSingapore100% (2)
- s41396 022 01245 4Document13 pagess41396 022 01245 4miralvaNo ratings yet
- Antimicrobial Activities of Leaves and Stem Bark Extracts of Buddleja SalviifoliaDocument8 pagesAntimicrobial Activities of Leaves and Stem Bark Extracts of Buddleja SalviifoliaPaul MatamaneNo ratings yet
- GscholarDocument5 pagesGscholarkikiNo ratings yet
- Diversity of Leaf Surface Fungi On The Medicinal Plant, Plectranthus Amboinicus Lour. and Its Antimicrobial Efficacy Against MTCC PathogensDocument6 pagesDiversity of Leaf Surface Fungi On The Medicinal Plant, Plectranthus Amboinicus Lour. and Its Antimicrobial Efficacy Against MTCC PathogensRanadive AnanthNo ratings yet
- The in Vitro Study of Antimicrobial Effect of Marigoldcalendula Officinalis Extract On Infectious MicroorganismsDocument5 pagesThe in Vitro Study of Antimicrobial Effect of Marigoldcalendula Officinalis Extract On Infectious MicroorganismsdssgssNo ratings yet
- EndophytesDocument19 pagesEndophytesAditya KanwalNo ratings yet
- Constantinidou53 BacterialcolonizationofthephyllosphereofnineteenplantspeciesandantimicrobialactivityoftheirleafsecondarymetabolitesagainstleafassociatedbacteriaDocument11 pagesConstantinidou53 BacterialcolonizationofthephyllosphereofnineteenplantspeciesandantimicrobialactivityoftheirleafsecondarymetabolitesagainstleafassociatedbacteriaindahdwirahNo ratings yet
- Endophytes To The Rescue of Plants!: Bhavdish N. JohriDocument2 pagesEndophytes To The Rescue of Plants!: Bhavdish N. JohriLuzAvNo ratings yet
- Vahling-Armstrong Et Al-2016-Plant PathologyDocument9 pagesVahling-Armstrong Et Al-2016-Plant PathologyJhon ReyesNo ratings yet
- Allelopathic Potential and HPTLC Analysis of Ipomoea Carnea PDFDocument5 pagesAllelopathic Potential and HPTLC Analysis of Ipomoea Carnea PDFSSR-IIJLS JournalNo ratings yet
- 1 s2.0 S0362028X22084447 MainDocument9 pages1 s2.0 S0362028X22084447 MainTanti 01No ratings yet
- Studies On Antibacterial Activity of Pithecellobium Dulce (Roxb.) Benth Against Food Pathogens - Gram Negative BacteriaDocument5 pagesStudies On Antibacterial Activity of Pithecellobium Dulce (Roxb.) Benth Against Food Pathogens - Gram Negative BacteriaDeepak ManogaranNo ratings yet
- Pathogenic FungiDocument2 pagesPathogenic FungiLeda Rodriguez PossoNo ratings yet
- Vitro AnuuDocument7 pagesVitro AnuusheikmoonchildNo ratings yet
- Powdery Mildew 2008 - Genetics and ResistanceDocument5 pagesPowdery Mildew 2008 - Genetics and ResistancenormanwillowNo ratings yet
- The Chaos of English PronunciationDocument5 pagesThe Chaos of English PronunciationMaría Regal CarballoNo ratings yet
- Bioterrorism Bioterrorism and Biological Warfare AgentsDocument10 pagesBioterrorism Bioterrorism and Biological Warfare AgentsDeng Navarro PasionNo ratings yet
- The Demon in The Freezer - Wikipedia, The Free EncyclopediaDocument4 pagesThe Demon in The Freezer - Wikipedia, The Free EncyclopediakibbyxdNo ratings yet
- Frac List of Plant Pathogenic Organisms Resistant To Disease Control AgentsDocument75 pagesFrac List of Plant Pathogenic Organisms Resistant To Disease Control AgentsGeraldNo ratings yet
- RubellaDocument49 pagesRubellarusdiNo ratings yet
- COVID 19 Genome Sequencing and DetectionDocument3 pagesCOVID 19 Genome Sequencing and DetectiongygyNo ratings yet
- E.Coli Coliforms Chromogenic Medium: CAT Nº: 1340Document2 pagesE.Coli Coliforms Chromogenic Medium: CAT Nº: 1340Juan Manuel Ramos ReyesNo ratings yet
- ChapterDocument82 pagesChapterScrewTheSchoolSystem100% (2)
- Mosquito ControlDocument284 pagesMosquito Controlrijade100% (1)
- Haad Exam 7Document22 pagesHaad Exam 7MallikarjunNo ratings yet
- FIP Cat TreatmentDocument14 pagesFIP Cat TreatmentRaihanNo ratings yet
- The Function of The PointsDocument175 pagesThe Function of The Pointsacuhome100% (2)
- Case Study: PharyngitisDocument3 pagesCase Study: PharyngitisRestian Rony SaragiNo ratings yet
- Assignment 1 Part 1 A1Document10 pagesAssignment 1 Part 1 A1garrenaNo ratings yet
- Edward JennerDocument5 pagesEdward JennerAnto FreistyawanNo ratings yet
- Sample Research Plan2Document9 pagesSample Research Plan2api-283862617No ratings yet
- Very Short Answer Questions: Microorganisms: Friend and Foe - S Chand Hops ClassDocument5 pagesVery Short Answer Questions: Microorganisms: Friend and Foe - S Chand Hops ClassShubhamNo ratings yet
- A Review of Traditional and Prospective Medicinal Applications of GarlicDocument10 pagesA Review of Traditional and Prospective Medicinal Applications of GarlicBrennan GarlandNo ratings yet
- Lecture Nursing Communicable Disease EpidemiologyDocument32 pagesLecture Nursing Communicable Disease EpidemiologyvolaaNo ratings yet
- Medical and Surgical NursingDocument4 pagesMedical and Surgical NursingCrystal Ann Monsale TadiamonNo ratings yet
- Health Care Waste Management of Funeral Homes in Cebu City - RRLDocument16 pagesHealth Care Waste Management of Funeral Homes in Cebu City - RRLDiane Sumalinog100% (1)
- Journal Predictive Factors For Lower Extremity Amputations in Diabetic Foot Infections - Dr. Samuel Jason Rolando Tua TobingDocument6 pagesJournal Predictive Factors For Lower Extremity Amputations in Diabetic Foot Infections - Dr. Samuel Jason Rolando Tua Tobingsamuel3101jasoNo ratings yet
- Trypanosoma Brucei: by Christian Jay N. BacantoDocument12 pagesTrypanosoma Brucei: by Christian Jay N. BacantoCHRISTIAN JAY BACANTONo ratings yet
- Blood Transfusion Reactions Revisited Dr. Edwin RodriguezDocument99 pagesBlood Transfusion Reactions Revisited Dr. Edwin RodriguezBeng AquinoNo ratings yet
- 4.vector and Water Borne-DiseasesDocument26 pages4.vector and Water Borne-DiseasesMuhammad AsifNo ratings yet
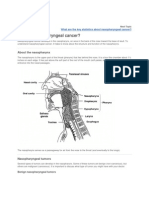
- Nasopharyngeal CancerDocument2 pagesNasopharyngeal CancerCharo FloresNo ratings yet
- Covid 19 Article - 2Document8 pagesCovid 19 Article - 2Kiruba KaranNo ratings yet
- Arte Sun AteDocument9 pagesArte Sun Atebaitur_rohmah8372No ratings yet