
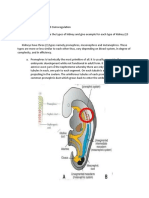
Anatomy and Physiology of The Reptile Renal System
Anatomy and Physiology of The Reptile Renal System
You might also like
- MODULE 2 Ichthyology - SECTION 5 Shark Anatomy and Dogfish Dissection PDFDocument27 pagesMODULE 2 Ichthyology - SECTION 5 Shark Anatomy and Dogfish Dissection PDFElishae SamonteNo ratings yet
- Cavitation RF Vaccumm EMSDocument11 pagesCavitation RF Vaccumm EMSLenny Rivero100% (2)
- Acute Flaccid Paralysis: Dr. Madoori SrinivasDocument53 pagesAcute Flaccid Paralysis: Dr. Madoori SrinivasdrravindermehetreyNo ratings yet
- Victoria Martens Autopsey 1484008212Document31 pagesVictoria Martens Autopsey 1484008212Anonymous Rbn0yhT100% (2)
- Simion Ana-MariaDocument8 pagesSimion Ana-MariaAna-Maria SimionNo ratings yet
- BIO 342 Comparative Vertebrate Anatomy Lecture Notes 10 - Urogenital SystemDocument4 pagesBIO 342 Comparative Vertebrate Anatomy Lecture Notes 10 - Urogenital SystemKatrina Lee GaganNo ratings yet
- Urogenital SystemDocument11 pagesUrogenital Systemchryan1989100% (2)
- Excretory System and OsmoregulationDocument10 pagesExcretory System and OsmoregulationShaira CogollodoNo ratings yet
- Dogfish Shark Dissection:: Squalus Acanthius)Document34 pagesDogfish Shark Dissection:: Squalus Acanthius)Bio SciencesNo ratings yet
- Amphibians: Introduction and EvolutionDocument5 pagesAmphibians: Introduction and Evolutionmurse101No ratings yet
- Ciclatory Systems of Different OrganismsDocument5 pagesCiclatory Systems of Different OrganismsSekhar ReddyNo ratings yet
- Research - 1Document23 pagesResearch - 1Osama ALGabriNo ratings yet
- Urinogenital System in EnglisDocument27 pagesUrinogenital System in EnglisYogaPradiptaNo ratings yet
- Sub-Phylum Cephalochordata (Lancelets) : ObjectivesDocument5 pagesSub-Phylum Cephalochordata (Lancelets) : ObjectivesMiftah RohmahNo ratings yet
- 1st PartDocument24 pages1st Partعزالدين الطيارNo ratings yet
- The Urogenital System: I. Vertebrate KidneysDocument5 pagesThe Urogenital System: I. Vertebrate KidneysSebastian SmytheNo ratings yet
- SquidDocument6 pagesSquidhaneenNo ratings yet
- Special Chordate CharacteriscticsDocument19 pagesSpecial Chordate CharacteriscticsKunica SurajNo ratings yet
- Biology of Ciona (Continuation)Document5 pagesBiology of Ciona (Continuation)Sulaiman OlanrewajuNo ratings yet
- Bird Kidney SystemDocument9 pagesBird Kidney SystemScribdTranslationsNo ratings yet
- AmphioxusDocument20 pagesAmphioxusmuhammadismailNo ratings yet
- Subphylum Cephalocordata: ClassificationDocument13 pagesSubphylum Cephalocordata: ClassificationMemona MumtazNo ratings yet
- Class ReptiliaDocument64 pagesClass ReptiliaMidnightNo ratings yet
- 463 - Fis 301Document9 pages463 - Fis 301aditasfadityaNo ratings yet
- Osmoregulation: Biology (../index - HTML) / Oc-Ph (Index - HTML) / OsmoregulationDocument7 pagesOsmoregulation: Biology (../index - HTML) / Oc-Ph (Index - HTML) / Osmoregulationdr_swaralipiNo ratings yet
- 3.swim Bladder PG MamDocument31 pages3.swim Bladder PG MamPrasun GhoshNo ratings yet
- Excretory System of InvertebratesDocument4 pagesExcretory System of InvertebratesHINA SHAHZADINo ratings yet
- Digestive System: Balanoglossus GigasDocument3 pagesDigestive System: Balanoglossus GigasJayashree RavishankarNo ratings yet
- Excretory System in InvertebratesDocument2 pagesExcretory System in Invertebratesihtishamulhaq1348No ratings yet
- Renal Anatomy and PhysiologyDocument15 pagesRenal Anatomy and PhysiologySapna JainNo ratings yet
- Phylum Annelida 1Document45 pagesPhylum Annelida 1b6330025No ratings yet
- ProtochordataDocument7 pagesProtochordataricoghofarNo ratings yet
- Nuevo Documento de TextoDocument9 pagesNuevo Documento de TextoyyyNo ratings yet
- Water Vascular System - Wikipedia, The Free Encyclopedia PDFDocument3 pagesWater Vascular System - Wikipedia, The Free Encyclopedia PDFDebarunAcharyaNo ratings yet
- Wyneken The Anatomy of Sea Turtle PDFDocument14 pagesWyneken The Anatomy of Sea Turtle PDFElis PerroneNo ratings yet
- Avian Renal System Clinical ImplicationsDocument19 pagesAvian Renal System Clinical ImplicationsAnais FaveroNo ratings yet
- The PolychaetesDocument3 pagesThe PolychaetesKiki_Amamanglon_3115No ratings yet
- Excretory PDFDocument104 pagesExcretory PDFgerman guazaNo ratings yet
- Full Download Book Essential Matlab For Engineers and Scientists PDFDocument22 pagesFull Download Book Essential Matlab For Engineers and Scientists PDFkimberly.davis912100% (25)
- Animal Kingdom 13Document17 pagesAnimal Kingdom 13Vicky KinsNo ratings yet
- 0 Bryozoa PDFDocument4 pages0 Bryozoa PDFMark Robert MagsinoNo ratings yet
- Lab 4 - Molluscs and AnnelidsDocument10 pagesLab 4 - Molluscs and AnnelidsNTA UGC-NETNo ratings yet
- Presentation 3Document24 pagesPresentation 3api-251518159No ratings yet
- Excretion of WaterDocument5 pagesExcretion of WaterKamrul Alam MasumNo ratings yet
- Hafeezullah: Subject:Anatomy Topic: Anatomy of Excretory System Roll No: 15 Semester: 1 RepeaterDocument10 pagesHafeezullah: Subject:Anatomy Topic: Anatomy of Excretory System Roll No: 15 Semester: 1 RepeaterMuhammad ShahzaibNo ratings yet
- How To Dissect A SharkDocument14 pagesHow To Dissect A SharkgregNo ratings yet
- Excretory System of FrogDocument3 pagesExcretory System of FrogSree Lekshmi V SNo ratings yet
- EarthwormDocument11 pagesEarthwormFabian GuadarramaNo ratings yet
- Respiring Atria Ventricle Aorta Pulmonary Artery: Digestion and ExcretionDocument5 pagesRespiring Atria Ventricle Aorta Pulmonary Artery: Digestion and Excretionkbarn389No ratings yet
- A Trouts Anatomy-WebDocument2 pagesA Trouts Anatomy-WebDiaconu DragosNo ratings yet
- M.Sc. II Sem. (Zoology) : Primitive Characters of DipnoiDocument11 pagesM.Sc. II Sem. (Zoology) : Primitive Characters of DipnoirajuNo ratings yet
- (JR., Cleveland P Hickman, Larry S Roberts, Allan OrgDocument58 pages(JR., Cleveland P Hickman, Larry S Roberts, Allan Orgalem010No ratings yet
- Questions and Their Ideal AnswersDocument17 pagesQuestions and Their Ideal AnswersHamzaShabaka100% (2)
- Biology of Balanoglossus (Continues)Document3 pagesBiology of Balanoglossus (Continues)Sulaiman OlanrewajuNo ratings yet
- Revision Notes: Excretory OrgansDocument4 pagesRevision Notes: Excretory Organskavitaruby1980No ratings yet
- Introduction To ChordataDocument203 pagesIntroduction To ChordataYhan Brotamonte BoneoNo ratings yet
- Human Excretory System - Grade 11 & 12Document19 pagesHuman Excretory System - Grade 11 & 12Ishita SinghNo ratings yet
- Zoology Assigment: 05: AcanthocephalaDocument6 pagesZoology Assigment: 05: AcanthocephalaLaiba ShNo ratings yet
- Lecture 06 - Cephalochordata and VertebrataDocument20 pagesLecture 06 - Cephalochordata and Vertebratamasif gondalNo ratings yet
- 0 Echinoderms-FinalDocument18 pages0 Echinoderms-FinalMark Robert MagsinoNo ratings yet
- Evolution of SpidersDocument15 pagesEvolution of SpidersHammad RazaNo ratings yet
- 2863-002 (Chương 3)Document47 pages2863-002 (Chương 3)Gia Bảo NguyễnNo ratings yet
- Nutrient Requirements of Rabbits, 2nd Revised Edition, 1977 - National Research CouncilDocument38 pagesNutrient Requirements of Rabbits, 2nd Revised Edition, 1977 - National Research CouncilEduardo LeónNo ratings yet
- Structure, Innervation and Response Properties of Integumentary Sensory Organs in CrocodiliansDocument14 pagesStructure, Innervation and Response Properties of Integumentary Sensory Organs in CrocodiliansEduardo LeónNo ratings yet
- Integumentary Sensory Organs in CrocodiliansDocument10 pagesIntegumentary Sensory Organs in CrocodiliansEduardo LeónNo ratings yet
- La Extinción Después de La Reactivación de La Memoria Del Miedo No Elimina La Renovación en RatasDocument7 pagesLa Extinción Después de La Reactivación de La Memoria Del Miedo No Elimina La Renovación en RatasEduardo LeónNo ratings yet
- The Use of Punishment For Behavior Modification in Animals PDFDocument4 pagesThe Use of Punishment For Behavior Modification in Animals PDFEduardo LeónNo ratings yet
- Examen Fisico General en ReptilesDocument37 pagesExamen Fisico General en ReptilesEduardo LeónNo ratings yet
- PleurodesisDocument6 pagesPleurodesisEduardo LeónNo ratings yet
- Afast TfastDocument7 pagesAfast TfastEduardo LeónNo ratings yet
- Core Stability - A Pilates WorkoutDocument26 pagesCore Stability - A Pilates WorkoutJelena VivodaNo ratings yet
- Overview of Food Microbiology: ObjectivesDocument35 pagesOverview of Food Microbiology: ObjectivesAshraf w50% (2)
- Users-Clinical-Cases 1607 Rev2 WebDocument46 pagesUsers-Clinical-Cases 1607 Rev2 WebJ.A. Lorenz AdlerNo ratings yet
- Assessment of Skin Integrity: Nursing ProcessDocument17 pagesAssessment of Skin Integrity: Nursing ProcessCONEY CHING SAINo ratings yet
- A Pattern Approach To Lymph Node Diagnosis - A. Leong (Springer, 2011) WWDocument432 pagesA Pattern Approach To Lymph Node Diagnosis - A. Leong (Springer, 2011) WWAdrian TirlaNo ratings yet
- 2019 AUG 7 Phe Epidemiological Update MeaslesDocument10 pages2019 AUG 7 Phe Epidemiological Update MeaslesJENIFFER NATALY ALVAREZ GALINDONo ratings yet
- Bohol Rabies Prevention & Eradication ProgramDocument42 pagesBohol Rabies Prevention & Eradication ProgramJaz MeNo ratings yet
- Body Fluids and Circulation Class 11 Notes CBSE Biology Chapter 18 (PDF)Document11 pagesBody Fluids and Circulation Class 11 Notes CBSE Biology Chapter 18 (PDF)Nilima Aparajita SahuNo ratings yet
- Nutritional Contribution of Meat and PoultryDocument14 pagesNutritional Contribution of Meat and Poultryank29jpleaseNo ratings yet
- Day 5 - Kuliah Heat StrokeDocument30 pagesDay 5 - Kuliah Heat StrokeMaha Satya Dwi PalgunaNo ratings yet
- Guide To Canadian Farmed SalmonDocument24 pagesGuide To Canadian Farmed SalmonKraft DinnerNo ratings yet
- Pemeriksaan Anemia SMP Darul MaarifDocument38 pagesPemeriksaan Anemia SMP Darul MaarifPuskesmas BanyuputihNo ratings yet
- Chapter 2 - Body CoordinationDocument13 pagesChapter 2 - Body CoordinationChai Wanzhen100% (1)
- Cancer - Rath, Matthias PDF (Pauling-Rath Therapy Protocol: Vitamin C and Lysine)Document84 pagesCancer - Rath, Matthias PDF (Pauling-Rath Therapy Protocol: Vitamin C and Lysine)Anonymous Jap77xvqPKNo ratings yet
- 2 Neurophysiology of Somatic Sensations 2012Document6 pages2 Neurophysiology of Somatic Sensations 2012Nephritery Joy Espiritu JimenezNo ratings yet
- L3 AP 101 Mock Questions Exam Guide V101.2Document24 pagesL3 AP 101 Mock Questions Exam Guide V101.2Sergiu TitcuNo ratings yet
- Quarter: 4 / Semester: 2 / WEEK: 4: Online Resource/sDocument17 pagesQuarter: 4 / Semester: 2 / WEEK: 4: Online Resource/sTristan Paul PagalananNo ratings yet
- Liver:: Anatomy The Abdomen Liver and Gall BladderDocument5 pagesLiver:: Anatomy The Abdomen Liver and Gall BladderWaeel AbdullatifNo ratings yet
- Raising Healthy Rabbits Under Primitive ConditionsDocument83 pagesRaising Healthy Rabbits Under Primitive ConditionsplutocowNo ratings yet
- Cocculus IndicusDocument29 pagesCocculus IndicusShah FaisalNo ratings yet
- What Is Malocclusion?Document24 pagesWhat Is Malocclusion?Bibek RajNo ratings yet
- 379 716 1 SMDocument10 pages379 716 1 SMRahmini RangkutiNo ratings yet
- Pregnancy Complication: Department of Gynaecology and ObstetricsDocument59 pagesPregnancy Complication: Department of Gynaecology and ObstetricssanjivdasNo ratings yet
- Lesson Plan: Achievement ObjectivesDocument4 pagesLesson Plan: Achievement ObjectivesAnonymous bStw5dNo ratings yet
- Nursing FundamentalsDocument41 pagesNursing FundamentalsEsther O. CorreaNo ratings yet
- Larvicidal Potency of MARIGOLD Stem andDocument24 pagesLarvicidal Potency of MARIGOLD Stem andAira Shayne ComeNo ratings yet
- A Guide To UCT's Third Tier Accommodation: Handbook 2.3Document14 pagesA Guide To UCT's Third Tier Accommodation: Handbook 2.3Mianda InstituteNo ratings yet

































































































Download as pdf or txt
You might also like
- MODULE 2 Ichthyology - SECTION 5 Shark Anatomy and Dogfish Dissection PDFDocument27 pagesMODULE 2 Ichthyology - SECTION 5 Shark Anatomy and Dogfish Dissection PDFElishae SamonteNo ratings yet
- Cavitation RF Vaccumm EMSDocument11 pagesCavitation RF Vaccumm EMSLenny Rivero100% (2)
- Acute Flaccid Paralysis: Dr. Madoori SrinivasDocument53 pagesAcute Flaccid Paralysis: Dr. Madoori SrinivasdrravindermehetreyNo ratings yet
- Victoria Martens Autopsey 1484008212Document31 pagesVictoria Martens Autopsey 1484008212Anonymous Rbn0yhT100% (2)
- Simion Ana-MariaDocument8 pagesSimion Ana-MariaAna-Maria SimionNo ratings yet
- BIO 342 Comparative Vertebrate Anatomy Lecture Notes 10 - Urogenital SystemDocument4 pagesBIO 342 Comparative Vertebrate Anatomy Lecture Notes 10 - Urogenital SystemKatrina Lee GaganNo ratings yet
- Urogenital SystemDocument11 pagesUrogenital Systemchryan1989100% (2)
- Excretory System and OsmoregulationDocument10 pagesExcretory System and OsmoregulationShaira CogollodoNo ratings yet
- Dogfish Shark Dissection:: Squalus Acanthius)Document34 pagesDogfish Shark Dissection:: Squalus Acanthius)Bio SciencesNo ratings yet
- Amphibians: Introduction and EvolutionDocument5 pagesAmphibians: Introduction and Evolutionmurse101No ratings yet
- Ciclatory Systems of Different OrganismsDocument5 pagesCiclatory Systems of Different OrganismsSekhar ReddyNo ratings yet
- Research - 1Document23 pagesResearch - 1Osama ALGabriNo ratings yet
- Urinogenital System in EnglisDocument27 pagesUrinogenital System in EnglisYogaPradiptaNo ratings yet
- Sub-Phylum Cephalochordata (Lancelets) : ObjectivesDocument5 pagesSub-Phylum Cephalochordata (Lancelets) : ObjectivesMiftah RohmahNo ratings yet
- 1st PartDocument24 pages1st Partعزالدين الطيارNo ratings yet
- The Urogenital System: I. Vertebrate KidneysDocument5 pagesThe Urogenital System: I. Vertebrate KidneysSebastian SmytheNo ratings yet
- SquidDocument6 pagesSquidhaneenNo ratings yet
- Special Chordate CharacteriscticsDocument19 pagesSpecial Chordate CharacteriscticsKunica SurajNo ratings yet
- Biology of Ciona (Continuation)Document5 pagesBiology of Ciona (Continuation)Sulaiman OlanrewajuNo ratings yet
- Bird Kidney SystemDocument9 pagesBird Kidney SystemScribdTranslationsNo ratings yet
- AmphioxusDocument20 pagesAmphioxusmuhammadismailNo ratings yet
- Subphylum Cephalocordata: ClassificationDocument13 pagesSubphylum Cephalocordata: ClassificationMemona MumtazNo ratings yet
- Class ReptiliaDocument64 pagesClass ReptiliaMidnightNo ratings yet
- 463 - Fis 301Document9 pages463 - Fis 301aditasfadityaNo ratings yet
- Osmoregulation: Biology (../index - HTML) / Oc-Ph (Index - HTML) / OsmoregulationDocument7 pagesOsmoregulation: Biology (../index - HTML) / Oc-Ph (Index - HTML) / Osmoregulationdr_swaralipiNo ratings yet
- 3.swim Bladder PG MamDocument31 pages3.swim Bladder PG MamPrasun GhoshNo ratings yet
- Excretory System of InvertebratesDocument4 pagesExcretory System of InvertebratesHINA SHAHZADINo ratings yet
- Digestive System: Balanoglossus GigasDocument3 pagesDigestive System: Balanoglossus GigasJayashree RavishankarNo ratings yet
- Excretory System in InvertebratesDocument2 pagesExcretory System in Invertebratesihtishamulhaq1348No ratings yet
- Renal Anatomy and PhysiologyDocument15 pagesRenal Anatomy and PhysiologySapna JainNo ratings yet
- Phylum Annelida 1Document45 pagesPhylum Annelida 1b6330025No ratings yet
- ProtochordataDocument7 pagesProtochordataricoghofarNo ratings yet
- Nuevo Documento de TextoDocument9 pagesNuevo Documento de TextoyyyNo ratings yet
- Water Vascular System - Wikipedia, The Free Encyclopedia PDFDocument3 pagesWater Vascular System - Wikipedia, The Free Encyclopedia PDFDebarunAcharyaNo ratings yet
- Wyneken The Anatomy of Sea Turtle PDFDocument14 pagesWyneken The Anatomy of Sea Turtle PDFElis PerroneNo ratings yet
- Avian Renal System Clinical ImplicationsDocument19 pagesAvian Renal System Clinical ImplicationsAnais FaveroNo ratings yet
- The PolychaetesDocument3 pagesThe PolychaetesKiki_Amamanglon_3115No ratings yet
- Excretory PDFDocument104 pagesExcretory PDFgerman guazaNo ratings yet
- Full Download Book Essential Matlab For Engineers and Scientists PDFDocument22 pagesFull Download Book Essential Matlab For Engineers and Scientists PDFkimberly.davis912100% (25)
- Animal Kingdom 13Document17 pagesAnimal Kingdom 13Vicky KinsNo ratings yet
- 0 Bryozoa PDFDocument4 pages0 Bryozoa PDFMark Robert MagsinoNo ratings yet
- Lab 4 - Molluscs and AnnelidsDocument10 pagesLab 4 - Molluscs and AnnelidsNTA UGC-NETNo ratings yet
- Presentation 3Document24 pagesPresentation 3api-251518159No ratings yet
- Excretion of WaterDocument5 pagesExcretion of WaterKamrul Alam MasumNo ratings yet
- Hafeezullah: Subject:Anatomy Topic: Anatomy of Excretory System Roll No: 15 Semester: 1 RepeaterDocument10 pagesHafeezullah: Subject:Anatomy Topic: Anatomy of Excretory System Roll No: 15 Semester: 1 RepeaterMuhammad ShahzaibNo ratings yet
- How To Dissect A SharkDocument14 pagesHow To Dissect A SharkgregNo ratings yet
- Excretory System of FrogDocument3 pagesExcretory System of FrogSree Lekshmi V SNo ratings yet
- EarthwormDocument11 pagesEarthwormFabian GuadarramaNo ratings yet
- Respiring Atria Ventricle Aorta Pulmonary Artery: Digestion and ExcretionDocument5 pagesRespiring Atria Ventricle Aorta Pulmonary Artery: Digestion and Excretionkbarn389No ratings yet
- A Trouts Anatomy-WebDocument2 pagesA Trouts Anatomy-WebDiaconu DragosNo ratings yet
- M.Sc. II Sem. (Zoology) : Primitive Characters of DipnoiDocument11 pagesM.Sc. II Sem. (Zoology) : Primitive Characters of DipnoirajuNo ratings yet
- (JR., Cleveland P Hickman, Larry S Roberts, Allan OrgDocument58 pages(JR., Cleveland P Hickman, Larry S Roberts, Allan Orgalem010No ratings yet
- Questions and Their Ideal AnswersDocument17 pagesQuestions and Their Ideal AnswersHamzaShabaka100% (2)
- Biology of Balanoglossus (Continues)Document3 pagesBiology of Balanoglossus (Continues)Sulaiman OlanrewajuNo ratings yet
- Revision Notes: Excretory OrgansDocument4 pagesRevision Notes: Excretory Organskavitaruby1980No ratings yet
- Introduction To ChordataDocument203 pagesIntroduction To ChordataYhan Brotamonte BoneoNo ratings yet
- Human Excretory System - Grade 11 & 12Document19 pagesHuman Excretory System - Grade 11 & 12Ishita SinghNo ratings yet
- Zoology Assigment: 05: AcanthocephalaDocument6 pagesZoology Assigment: 05: AcanthocephalaLaiba ShNo ratings yet
- Lecture 06 - Cephalochordata and VertebrataDocument20 pagesLecture 06 - Cephalochordata and Vertebratamasif gondalNo ratings yet
- 0 Echinoderms-FinalDocument18 pages0 Echinoderms-FinalMark Robert MagsinoNo ratings yet
- Evolution of SpidersDocument15 pagesEvolution of SpidersHammad RazaNo ratings yet
- 2863-002 (Chương 3)Document47 pages2863-002 (Chương 3)Gia Bảo NguyễnNo ratings yet
- Nutrient Requirements of Rabbits, 2nd Revised Edition, 1977 - National Research CouncilDocument38 pagesNutrient Requirements of Rabbits, 2nd Revised Edition, 1977 - National Research CouncilEduardo LeónNo ratings yet
- Structure, Innervation and Response Properties of Integumentary Sensory Organs in CrocodiliansDocument14 pagesStructure, Innervation and Response Properties of Integumentary Sensory Organs in CrocodiliansEduardo LeónNo ratings yet
- Integumentary Sensory Organs in CrocodiliansDocument10 pagesIntegumentary Sensory Organs in CrocodiliansEduardo LeónNo ratings yet
- La Extinción Después de La Reactivación de La Memoria Del Miedo No Elimina La Renovación en RatasDocument7 pagesLa Extinción Después de La Reactivación de La Memoria Del Miedo No Elimina La Renovación en RatasEduardo LeónNo ratings yet
- The Use of Punishment For Behavior Modification in Animals PDFDocument4 pagesThe Use of Punishment For Behavior Modification in Animals PDFEduardo LeónNo ratings yet
- Examen Fisico General en ReptilesDocument37 pagesExamen Fisico General en ReptilesEduardo LeónNo ratings yet
- PleurodesisDocument6 pagesPleurodesisEduardo LeónNo ratings yet
- Afast TfastDocument7 pagesAfast TfastEduardo LeónNo ratings yet
- Core Stability - A Pilates WorkoutDocument26 pagesCore Stability - A Pilates WorkoutJelena VivodaNo ratings yet
- Overview of Food Microbiology: ObjectivesDocument35 pagesOverview of Food Microbiology: ObjectivesAshraf w50% (2)
- Users-Clinical-Cases 1607 Rev2 WebDocument46 pagesUsers-Clinical-Cases 1607 Rev2 WebJ.A. Lorenz AdlerNo ratings yet
- Assessment of Skin Integrity: Nursing ProcessDocument17 pagesAssessment of Skin Integrity: Nursing ProcessCONEY CHING SAINo ratings yet
- A Pattern Approach To Lymph Node Diagnosis - A. Leong (Springer, 2011) WWDocument432 pagesA Pattern Approach To Lymph Node Diagnosis - A. Leong (Springer, 2011) WWAdrian TirlaNo ratings yet
- 2019 AUG 7 Phe Epidemiological Update MeaslesDocument10 pages2019 AUG 7 Phe Epidemiological Update MeaslesJENIFFER NATALY ALVAREZ GALINDONo ratings yet
- Bohol Rabies Prevention & Eradication ProgramDocument42 pagesBohol Rabies Prevention & Eradication ProgramJaz MeNo ratings yet
- Body Fluids and Circulation Class 11 Notes CBSE Biology Chapter 18 (PDF)Document11 pagesBody Fluids and Circulation Class 11 Notes CBSE Biology Chapter 18 (PDF)Nilima Aparajita SahuNo ratings yet
- Nutritional Contribution of Meat and PoultryDocument14 pagesNutritional Contribution of Meat and Poultryank29jpleaseNo ratings yet
- Day 5 - Kuliah Heat StrokeDocument30 pagesDay 5 - Kuliah Heat StrokeMaha Satya Dwi PalgunaNo ratings yet
- Guide To Canadian Farmed SalmonDocument24 pagesGuide To Canadian Farmed SalmonKraft DinnerNo ratings yet
- Pemeriksaan Anemia SMP Darul MaarifDocument38 pagesPemeriksaan Anemia SMP Darul MaarifPuskesmas BanyuputihNo ratings yet
- Chapter 2 - Body CoordinationDocument13 pagesChapter 2 - Body CoordinationChai Wanzhen100% (1)
- Cancer - Rath, Matthias PDF (Pauling-Rath Therapy Protocol: Vitamin C and Lysine)Document84 pagesCancer - Rath, Matthias PDF (Pauling-Rath Therapy Protocol: Vitamin C and Lysine)Anonymous Jap77xvqPKNo ratings yet
- 2 Neurophysiology of Somatic Sensations 2012Document6 pages2 Neurophysiology of Somatic Sensations 2012Nephritery Joy Espiritu JimenezNo ratings yet
- L3 AP 101 Mock Questions Exam Guide V101.2Document24 pagesL3 AP 101 Mock Questions Exam Guide V101.2Sergiu TitcuNo ratings yet
- Quarter: 4 / Semester: 2 / WEEK: 4: Online Resource/sDocument17 pagesQuarter: 4 / Semester: 2 / WEEK: 4: Online Resource/sTristan Paul PagalananNo ratings yet
- Liver:: Anatomy The Abdomen Liver and Gall BladderDocument5 pagesLiver:: Anatomy The Abdomen Liver and Gall BladderWaeel AbdullatifNo ratings yet
- Raising Healthy Rabbits Under Primitive ConditionsDocument83 pagesRaising Healthy Rabbits Under Primitive ConditionsplutocowNo ratings yet
- Cocculus IndicusDocument29 pagesCocculus IndicusShah FaisalNo ratings yet
- What Is Malocclusion?Document24 pagesWhat Is Malocclusion?Bibek RajNo ratings yet
- 379 716 1 SMDocument10 pages379 716 1 SMRahmini RangkutiNo ratings yet
- Pregnancy Complication: Department of Gynaecology and ObstetricsDocument59 pagesPregnancy Complication: Department of Gynaecology and ObstetricssanjivdasNo ratings yet
- Lesson Plan: Achievement ObjectivesDocument4 pagesLesson Plan: Achievement ObjectivesAnonymous bStw5dNo ratings yet
- Nursing FundamentalsDocument41 pagesNursing FundamentalsEsther O. CorreaNo ratings yet
- Larvicidal Potency of MARIGOLD Stem andDocument24 pagesLarvicidal Potency of MARIGOLD Stem andAira Shayne ComeNo ratings yet
- A Guide To UCT's Third Tier Accommodation: Handbook 2.3Document14 pagesA Guide To UCT's Third Tier Accommodation: Handbook 2.3Mianda InstituteNo ratings yet