
Role For Ferritin in Hematopoiesis and The Immune System
Role For Ferritin in Hematopoiesis and The Immune System
You might also like
- Biology 1A-Cell Motility Results and ConclusionsDocument6 pagesBiology 1A-Cell Motility Results and ConclusionsAna Maria100% (2)
- Ipe BrochureDocument2 pagesIpe Brochureapi-253709303No ratings yet
- Chapter 33 Guyton and HallDocument4 pagesChapter 33 Guyton and Hallg_komolafeNo ratings yet
- Ferritin and IsoferitinsDocument25 pagesFerritin and IsoferitinsAlin AlecuNo ratings yet
- Feritin - WIREs Nanomed Nanobiotechnol - 2013 - FanDocument12 pagesFeritin - WIREs Nanomed Nanobiotechnol - 2013 - FanAleksandar KitanovskiNo ratings yet
- Perforin and Granzymes: Function, Dysfunction and Human PathologyDocument13 pagesPerforin and Granzymes: Function, Dysfunction and Human Pathologyhoangphuong08101992No ratings yet
- Matrix Metalloproteinases and Their Inhibitors in ConnectiveDocument10 pagesMatrix Metalloproteinases and Their Inhibitors in ConnectiveBayu Anggariksa Alap AbdonNo ratings yet
- Serum Ferritin - Past, Present and FutureDocument21 pagesSerum Ferritin - Past, Present and FutureAleksandar KitanovskiNo ratings yet
- Production of Growth Factors, Cytokines, and ChemokinesDocument16 pagesProduction of Growth Factors, Cytokines, and ChemokinesNina TrươngNo ratings yet
- Gut Microbiota Sustains Hematopoiesis - BloodDocument3 pagesGut Microbiota Sustains Hematopoiesis - BloodFlávia UchôaNo ratings yet
- Perforin and Granzymes: Function, Dysfunction and Human PathologyDocument13 pagesPerforin and Granzymes: Function, Dysfunction and Human Pathologybca abcNo ratings yet
- Allergology InternationalDocument10 pagesAllergology InternationalaliNo ratings yet
- Haematopoiesis and Red Blood Cells 2013 Surgery OxfordDocument6 pagesHaematopoiesis and Red Blood Cells 2013 Surgery OxfordSanjaya SenevirathneNo ratings yet
- Leukocyte Platelet-Rich Fibrin (L-PRF), A New Biomembrane Useful in Tissue Repair: Basic Science and Literature ReviewDocument10 pagesLeukocyte Platelet-Rich Fibrin (L-PRF), A New Biomembrane Useful in Tissue Repair: Basic Science and Literature ReviewHtooNo ratings yet
- Impact of Chronic and Acute Inflammation On Extra - and Intracellular Iron Homeostasis (Ross, 2017)Document7 pagesImpact of Chronic and Acute Inflammation On Extra - and Intracellular Iron Homeostasis (Ross, 2017)Maria Luisa Sibajas ÁvilaNo ratings yet
- Molecular of Irhom.1Document14 pagesMolecular of Irhom.1MARIA DE FATIMA TEIXEIRA GORDILLONo ratings yet
- This Month S Highlights.38Document3 pagesThis Month S Highlights.38Arista RachmaNo ratings yet
- Eosinophils: Structure and Peter F Weller FunctionsDocument6 pagesEosinophils: Structure and Peter F Weller Functionsfernandaneves30No ratings yet
- Crit Model Anemia InflamationDocument3 pagesCrit Model Anemia Inflamationiris1088No ratings yet
- Evaluation of The Inhibitory Effect of Triphala On PMN-Type Matrix Metalloproteinase (MMP-9)Document6 pagesEvaluation of The Inhibitory Effect of Triphala On PMN-Type Matrix Metalloproteinase (MMP-9)Shankar arumugamNo ratings yet
- Ferroptosis and Parkinson's DiseaseDocument11 pagesFerroptosis and Parkinson's DiseaseParth RanaNo ratings yet
- 2007-Effects of TGF-beta2, BMP-4, and Gremlin in The Trabecular Meshwork Implications For GlaucomaDocument10 pages2007-Effects of TGF-beta2, BMP-4, and Gremlin in The Trabecular Meshwork Implications For GlaucomaReza NaghibiNo ratings yet
- Basic Concepts OF THE Mechanism OF Action OF Peptide HormonesDocument15 pagesBasic Concepts OF THE Mechanism OF Action OF Peptide Hormonesjuliana leonNo ratings yet
- Ferroptosis, A Novel Pharmacological Mechanism of Anti-Cancer DrugsDocument10 pagesFerroptosis, A Novel Pharmacological Mechanism of Anti-Cancer DrugsRob BordenNo ratings yet
- Erythropoietin Surprises. An Immune SagaDocument2 pagesErythropoietin Surprises. An Immune SagalmvalliNo ratings yet
- Anemia DLM Tinjauan PK Kuliah UmsDocument51 pagesAnemia DLM Tinjauan PK Kuliah UmsZammira MutiaNo ratings yet
- Emerging and Dynamic Biomedical Uses of Ferritin: PharmaceuticalsDocument18 pagesEmerging and Dynamic Biomedical Uses of Ferritin: PharmaceuticalsCristina CbNo ratings yet
- 10.1038@s41573 020 0063 yDocument13 pages10.1038@s41573 020 0063 ySpoti PremiNo ratings yet
- Eosinofilos Factores de Transcripcion en Su Desarrollo Como Blanco Terapeutico Frontiers Medicine 2017Document6 pagesEosinofilos Factores de Transcripcion en Su Desarrollo Como Blanco Terapeutico Frontiers Medicine 2017Gustavo GomezNo ratings yet
- Applications of Platelet Rich Plasma Fo PDFDocument6 pagesApplications of Platelet Rich Plasma Fo PDFRamona MateiNo ratings yet
- Applications of Platelet Rich Plasma PDFDocument6 pagesApplications of Platelet Rich Plasma PDFRamona MateiNo ratings yet
- Article Isoenzymes Clinical Diagnosis: SpecialDocument10 pagesArticle Isoenzymes Clinical Diagnosis: Specialanmol bandekarNo ratings yet
- Zinc Is A Novel Intracellular Second MessengerDocument9 pagesZinc Is A Novel Intracellular Second MessengerIdyah Hadiyanti A.No ratings yet
- 1 s2.0 S0006291X14003982 MainDocument5 pages1 s2.0 S0006291X14003982 Mainltthao.34.2023No ratings yet
- Ijms 25 00579 v2Document15 pagesIjms 25 00579 v2silviacas11No ratings yet
- Erythroferrone Structure, Function, and PhysiologyDocument24 pagesErythroferrone Structure, Function, and PhysiologyNia AvellanedaNo ratings yet
- Okuda JP03Document10 pagesOkuda JP03RohanGarudNo ratings yet
- The L PRF MembraneDocument11 pagesThe L PRF Membraneanh le ducNo ratings yet
- Basic Mechanisms and Regulation of Fibrinolysis: C.Longstaff Andk - KolevDocument8 pagesBasic Mechanisms and Regulation of Fibrinolysis: C.Longstaff Andk - Kolevmastmulla921No ratings yet
- Bond and Bulow - 2013 - Chimeric Genes-ProteinsDocument2 pagesBond and Bulow - 2013 - Chimeric Genes-ProteinsIsabella FernandesNo ratings yet
- Andi Muh. Octavian Pratama Et Anwar LewaDocument8 pagesAndi Muh. Octavian Pratama Et Anwar Lewaoctavian pratamaNo ratings yet
- Protein As MicromoleculesDocument7 pagesProtein As MicromoleculesPrincess JacintoNo ratings yet
- Andi Muh. Octavian Pratama Et Anwar Lewa-1Document9 pagesAndi Muh. Octavian Pratama Et Anwar Lewa-1octavian pratamaNo ratings yet
- Amphotericin B Inhibits Entry of Leishmania Donovani Into Primary MacrophagesDocument5 pagesAmphotericin B Inhibits Entry of Leishmania Donovani Into Primary Macrophagesyuvrajsingh3No ratings yet
- Ferritin Package InsertDocument2 pagesFerritin Package Insertluisoft88No ratings yet
- 04 - Iron Homeostasis in Host DefenceDocument11 pages04 - Iron Homeostasis in Host DefenceMatheus SouzaNo ratings yet
- 85471Document7 pages85471Vinnie Juliana YonatanNo ratings yet
- Enzimologia KanekoDocument28 pagesEnzimologia KanekoKatty Urbano OrdoñezNo ratings yet
- Review Article: Antioxidant Functions of Phytic AcidDocument9 pagesReview Article: Antioxidant Functions of Phytic AcidPhạm Nguyễn Minh ThuNo ratings yet
- Biochem Lec M-3Document13 pagesBiochem Lec M-3GE SalenNo ratings yet
- A Review On ErythropoietinDocument14 pagesA Review On ErythropoietinZulfkar Latief QadrieNo ratings yet
- The Chronobiology of Human Cytokines ProductionDocument15 pagesThe Chronobiology of Human Cytokines Productionيوميات صيدلانية pharmacist diariesNo ratings yet
- A New Portrait of Constitutive HeterochromatinDocument19 pagesA New Portrait of Constitutive HeterochromatinannyoryaNo ratings yet
- Triad Nihms 475933Document44 pagesTriad Nihms 475933poojaNo ratings yet
- Messengers From The MicrobiotaDocument3 pagesMessengers From The MicrobiotaDra. Ianna CarmoNo ratings yet
- Man Nello 2005Document15 pagesMan Nello 2005edelinNo ratings yet
- Ferroptosis: A Unique Form of Iron-Dependent Regulated Cell Death and Its Role in Different DiseasesDocument15 pagesFerroptosis: A Unique Form of Iron-Dependent Regulated Cell Death and Its Role in Different DiseasesSnehasis JanaNo ratings yet
- Virulence properties of the Legionella pneumophilaDocument12 pagesVirulence properties of the Legionella pneumophilaGuillermo RodríguezNo ratings yet
- The Role of Hepcidin in Iron MetabolismDocument10 pagesThe Role of Hepcidin in Iron MetabolismTatiana JalbaNo ratings yet
- Ferroptosis of Macrophages Facilitates Bone Loss in Apical Periodontitis Via NRF2 - FSP1 - ROS PathwayDocument14 pagesFerroptosis of Macrophages Facilitates Bone Loss in Apical Periodontitis Via NRF2 - FSP1 - ROS Pathwayratna saraswatiNo ratings yet
- Specs 5FE GBDocument5 pagesSpecs 5FE GBkemitaNo ratings yet
- Topnotch Surgery Supplement Handout UPDATED April 2016 PDFDocument83 pagesTopnotch Surgery Supplement Handout UPDATED April 2016 PDFDre ValdezNo ratings yet
- Susu HipoalergenikDocument14 pagesSusu HipoalergenikStella NoviaNo ratings yet
- Tumor Angiogenesis: Causes, Consequences, Challenges and OpportunitiesDocument26 pagesTumor Angiogenesis: Causes, Consequences, Challenges and OpportunitiesStella NoviaNo ratings yet
- Abnormal Vitamin Status in Childhood LeukemiaDocument8 pagesAbnormal Vitamin Status in Childhood LeukemiaStella NoviaNo ratings yet
- Effects of Vitamin B6 Metabolism On Oncogenesis, Tumor Progression and Therapeutic ResponsesDocument10 pagesEffects of Vitamin B6 Metabolism On Oncogenesis, Tumor Progression and Therapeutic ResponsesStella NoviaNo ratings yet
- Role of Central Vagal 5-HT Receptors in Gastrointestinal Physiology and PathophysiologyDocument10 pagesRole of Central Vagal 5-HT Receptors in Gastrointestinal Physiology and PathophysiologyStella NoviaNo ratings yet
- Free PPT Templates: Insert The Title of Your Presentation HereDocument3 pagesFree PPT Templates: Insert The Title of Your Presentation HereStella NoviaNo ratings yet
- Cancer Statistics, 2008Document26 pagesCancer Statistics, 2008Stella NoviaNo ratings yet
- Research ArticleDocument9 pagesResearch ArticleStella NoviaNo ratings yet
- Agenda Postgradmededu 120920 PDFDocument2 pagesAgenda Postgradmededu 120920 PDFStella NoviaNo ratings yet
- New England Journal Medicine: The ofDocument11 pagesNew England Journal Medicine: The ofStella NoviaNo ratings yet
- Nejmoa1911998 PDFDocument11 pagesNejmoa1911998 PDFStella NoviaNo ratings yet
- Free PPT Templates: PandemicDocument48 pagesFree PPT Templates: PandemicStella NoviaNo ratings yet
- Were The Babies Switched - Answer Sheet 2023 - GalvinDocument5 pagesWere The Babies Switched - Answer Sheet 2023 - Galvin25210580No ratings yet
- Reviewer (Abpsych)Document7 pagesReviewer (Abpsych)Joana Vivien Caraan100% (2)
- 2010-11 partA-F eDocument41 pages2010-11 partA-F eAltunc Suleyman GezerlerNo ratings yet
- Full Download Essentials of Physical Anthropology 10th Edition Jurmain Solutions ManualDocument35 pagesFull Download Essentials of Physical Anthropology 10th Edition Jurmain Solutions Manualudalbornhockeyspl2100% (31)
- Rheumatoid Arthritis: Pathophysiology PathophysiologyDocument34 pagesRheumatoid Arthritis: Pathophysiology PathophysiologyOmair RiazNo ratings yet
- Hormone 1Document14 pagesHormone 1IMDCBiochemNo ratings yet
- Spindle Cell Sarcomas: Cyril FisherDocument24 pagesSpindle Cell Sarcomas: Cyril FisherThierry GrimaNo ratings yet
- 1) Class 12th Biology Complete Unacademy BookDocument538 pages1) Class 12th Biology Complete Unacademy Bookkanikadudeja.07No ratings yet
- Biology Unit 3 Revision Year 8Document3 pagesBiology Unit 3 Revision Year 8Alan klnNo ratings yet
- PDF Diagnosis and Management of Mitochondrial Disorders Michelangelo Mancuso Ebook Full ChapterDocument53 pagesPDF Diagnosis and Management of Mitochondrial Disorders Michelangelo Mancuso Ebook Full Chapteredward.goodwin495100% (4)
- Detection of Malarial Parasite in Blood Using Image ProcessingDocument3 pagesDetection of Malarial Parasite in Blood Using Image ProcessingelectricalengineerNo ratings yet
- TA Brown Chapter 2 NotesDocument7 pagesTA Brown Chapter 2 Notesom prakashNo ratings yet
- File 1434571027Document1 pageFile 1434571027Guneyden GuneydenNo ratings yet
- Divine Intervention Episode 17 Diseases of The Pediatric Population Part 1Document14 pagesDivine Intervention Episode 17 Diseases of The Pediatric Population Part 1Swisskelly1No ratings yet
- Research Triangle Regions Life Sciences Directory 2012Document50 pagesResearch Triangle Regions Life Sciences Directory 2012hitexNo ratings yet
- The Immune System: HistoryDocument4 pagesThe Immune System: HistoryleesclassroomNo ratings yet
- A Project Proposal On Case Study and Management of A Type 1 Diabetes Mellitus ClientDocument6 pagesA Project Proposal On Case Study and Management of A Type 1 Diabetes Mellitus ClientMichael Kivumbi100% (3)
- An Update On Therapeutic Repurposing Strategies For COVID-19Document7 pagesAn Update On Therapeutic Repurposing Strategies For COVID-19Prakash KatakamNo ratings yet
- Listening Sample Test 1 Audio ScriptDocument11 pagesListening Sample Test 1 Audio ScriptCCCAXANo ratings yet
- Principles of Drug Addiction Treatment A Research Based Guide Third EditionDocument67 pagesPrinciples of Drug Addiction Treatment A Research Based Guide Third EditionКонстантин Крахмалев100% (1)
- Ankylosing Spondylitis (Marie-Strümpell Disease/ Bechterew's Disease)Document39 pagesAnkylosing Spondylitis (Marie-Strümpell Disease/ Bechterew's Disease)MoineeNo ratings yet
- Mod 2 Comp 13 - Demonstrate Knowledge of Electrical Safety and Safe Working Practices For Electrical WorkersDocument9 pagesMod 2 Comp 13 - Demonstrate Knowledge of Electrical Safety and Safe Working Practices For Electrical WorkersSmr OnlyNo ratings yet
- Biology - Form 2 Scheme of Work - Term 1: Week Topic Syllabus Objectives Number OF Sessions Activities/LabsDocument2 pagesBiology - Form 2 Scheme of Work - Term 1: Week Topic Syllabus Objectives Number OF Sessions Activities/LabsramjdNo ratings yet
- Juvenile Nasopharyngeal AngiofibromaDocument28 pagesJuvenile Nasopharyngeal AngiofibromamaNo ratings yet
- Health Benefits of Coconut Milk23Document6 pagesHealth Benefits of Coconut Milk23Gerson Paul BangoyNo ratings yet
- Premalignant LesionsDocument76 pagesPremalignant LesionsPrima D Andri100% (1)
- Small Animal OncologyDocument103 pagesSmall Animal Oncologyvet53No ratings yet
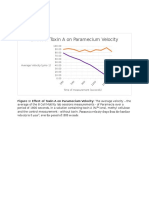
































































































Download as pdf or txt
You might also like
- Biology 1A-Cell Motility Results and ConclusionsDocument6 pagesBiology 1A-Cell Motility Results and ConclusionsAna Maria100% (2)
- Ipe BrochureDocument2 pagesIpe Brochureapi-253709303No ratings yet
- Chapter 33 Guyton and HallDocument4 pagesChapter 33 Guyton and Hallg_komolafeNo ratings yet
- Ferritin and IsoferitinsDocument25 pagesFerritin and IsoferitinsAlin AlecuNo ratings yet
- Feritin - WIREs Nanomed Nanobiotechnol - 2013 - FanDocument12 pagesFeritin - WIREs Nanomed Nanobiotechnol - 2013 - FanAleksandar KitanovskiNo ratings yet
- Perforin and Granzymes: Function, Dysfunction and Human PathologyDocument13 pagesPerforin and Granzymes: Function, Dysfunction and Human Pathologyhoangphuong08101992No ratings yet
- Matrix Metalloproteinases and Their Inhibitors in ConnectiveDocument10 pagesMatrix Metalloproteinases and Their Inhibitors in ConnectiveBayu Anggariksa Alap AbdonNo ratings yet
- Serum Ferritin - Past, Present and FutureDocument21 pagesSerum Ferritin - Past, Present and FutureAleksandar KitanovskiNo ratings yet
- Production of Growth Factors, Cytokines, and ChemokinesDocument16 pagesProduction of Growth Factors, Cytokines, and ChemokinesNina TrươngNo ratings yet
- Gut Microbiota Sustains Hematopoiesis - BloodDocument3 pagesGut Microbiota Sustains Hematopoiesis - BloodFlávia UchôaNo ratings yet
- Perforin and Granzymes: Function, Dysfunction and Human PathologyDocument13 pagesPerforin and Granzymes: Function, Dysfunction and Human Pathologybca abcNo ratings yet
- Allergology InternationalDocument10 pagesAllergology InternationalaliNo ratings yet
- Haematopoiesis and Red Blood Cells 2013 Surgery OxfordDocument6 pagesHaematopoiesis and Red Blood Cells 2013 Surgery OxfordSanjaya SenevirathneNo ratings yet
- Leukocyte Platelet-Rich Fibrin (L-PRF), A New Biomembrane Useful in Tissue Repair: Basic Science and Literature ReviewDocument10 pagesLeukocyte Platelet-Rich Fibrin (L-PRF), A New Biomembrane Useful in Tissue Repair: Basic Science and Literature ReviewHtooNo ratings yet
- Impact of Chronic and Acute Inflammation On Extra - and Intracellular Iron Homeostasis (Ross, 2017)Document7 pagesImpact of Chronic and Acute Inflammation On Extra - and Intracellular Iron Homeostasis (Ross, 2017)Maria Luisa Sibajas ÁvilaNo ratings yet
- Molecular of Irhom.1Document14 pagesMolecular of Irhom.1MARIA DE FATIMA TEIXEIRA GORDILLONo ratings yet
- This Month S Highlights.38Document3 pagesThis Month S Highlights.38Arista RachmaNo ratings yet
- Eosinophils: Structure and Peter F Weller FunctionsDocument6 pagesEosinophils: Structure and Peter F Weller Functionsfernandaneves30No ratings yet
- Crit Model Anemia InflamationDocument3 pagesCrit Model Anemia Inflamationiris1088No ratings yet
- Evaluation of The Inhibitory Effect of Triphala On PMN-Type Matrix Metalloproteinase (MMP-9)Document6 pagesEvaluation of The Inhibitory Effect of Triphala On PMN-Type Matrix Metalloproteinase (MMP-9)Shankar arumugamNo ratings yet
- Ferroptosis and Parkinson's DiseaseDocument11 pagesFerroptosis and Parkinson's DiseaseParth RanaNo ratings yet
- 2007-Effects of TGF-beta2, BMP-4, and Gremlin in The Trabecular Meshwork Implications For GlaucomaDocument10 pages2007-Effects of TGF-beta2, BMP-4, and Gremlin in The Trabecular Meshwork Implications For GlaucomaReza NaghibiNo ratings yet
- Basic Concepts OF THE Mechanism OF Action OF Peptide HormonesDocument15 pagesBasic Concepts OF THE Mechanism OF Action OF Peptide Hormonesjuliana leonNo ratings yet
- Ferroptosis, A Novel Pharmacological Mechanism of Anti-Cancer DrugsDocument10 pagesFerroptosis, A Novel Pharmacological Mechanism of Anti-Cancer DrugsRob BordenNo ratings yet
- Erythropoietin Surprises. An Immune SagaDocument2 pagesErythropoietin Surprises. An Immune SagalmvalliNo ratings yet
- Anemia DLM Tinjauan PK Kuliah UmsDocument51 pagesAnemia DLM Tinjauan PK Kuliah UmsZammira MutiaNo ratings yet
- Emerging and Dynamic Biomedical Uses of Ferritin: PharmaceuticalsDocument18 pagesEmerging and Dynamic Biomedical Uses of Ferritin: PharmaceuticalsCristina CbNo ratings yet
- 10.1038@s41573 020 0063 yDocument13 pages10.1038@s41573 020 0063 ySpoti PremiNo ratings yet
- Eosinofilos Factores de Transcripcion en Su Desarrollo Como Blanco Terapeutico Frontiers Medicine 2017Document6 pagesEosinofilos Factores de Transcripcion en Su Desarrollo Como Blanco Terapeutico Frontiers Medicine 2017Gustavo GomezNo ratings yet
- Applications of Platelet Rich Plasma Fo PDFDocument6 pagesApplications of Platelet Rich Plasma Fo PDFRamona MateiNo ratings yet
- Applications of Platelet Rich Plasma PDFDocument6 pagesApplications of Platelet Rich Plasma PDFRamona MateiNo ratings yet
- Article Isoenzymes Clinical Diagnosis: SpecialDocument10 pagesArticle Isoenzymes Clinical Diagnosis: Specialanmol bandekarNo ratings yet
- Zinc Is A Novel Intracellular Second MessengerDocument9 pagesZinc Is A Novel Intracellular Second MessengerIdyah Hadiyanti A.No ratings yet
- 1 s2.0 S0006291X14003982 MainDocument5 pages1 s2.0 S0006291X14003982 Mainltthao.34.2023No ratings yet
- Ijms 25 00579 v2Document15 pagesIjms 25 00579 v2silviacas11No ratings yet
- Erythroferrone Structure, Function, and PhysiologyDocument24 pagesErythroferrone Structure, Function, and PhysiologyNia AvellanedaNo ratings yet
- Okuda JP03Document10 pagesOkuda JP03RohanGarudNo ratings yet
- The L PRF MembraneDocument11 pagesThe L PRF Membraneanh le ducNo ratings yet
- Basic Mechanisms and Regulation of Fibrinolysis: C.Longstaff Andk - KolevDocument8 pagesBasic Mechanisms and Regulation of Fibrinolysis: C.Longstaff Andk - Kolevmastmulla921No ratings yet
- Bond and Bulow - 2013 - Chimeric Genes-ProteinsDocument2 pagesBond and Bulow - 2013 - Chimeric Genes-ProteinsIsabella FernandesNo ratings yet
- Andi Muh. Octavian Pratama Et Anwar LewaDocument8 pagesAndi Muh. Octavian Pratama Et Anwar Lewaoctavian pratamaNo ratings yet
- Protein As MicromoleculesDocument7 pagesProtein As MicromoleculesPrincess JacintoNo ratings yet
- Andi Muh. Octavian Pratama Et Anwar Lewa-1Document9 pagesAndi Muh. Octavian Pratama Et Anwar Lewa-1octavian pratamaNo ratings yet
- Amphotericin B Inhibits Entry of Leishmania Donovani Into Primary MacrophagesDocument5 pagesAmphotericin B Inhibits Entry of Leishmania Donovani Into Primary Macrophagesyuvrajsingh3No ratings yet
- Ferritin Package InsertDocument2 pagesFerritin Package Insertluisoft88No ratings yet
- 04 - Iron Homeostasis in Host DefenceDocument11 pages04 - Iron Homeostasis in Host DefenceMatheus SouzaNo ratings yet
- 85471Document7 pages85471Vinnie Juliana YonatanNo ratings yet
- Enzimologia KanekoDocument28 pagesEnzimologia KanekoKatty Urbano OrdoñezNo ratings yet
- Review Article: Antioxidant Functions of Phytic AcidDocument9 pagesReview Article: Antioxidant Functions of Phytic AcidPhạm Nguyễn Minh ThuNo ratings yet
- Biochem Lec M-3Document13 pagesBiochem Lec M-3GE SalenNo ratings yet
- A Review On ErythropoietinDocument14 pagesA Review On ErythropoietinZulfkar Latief QadrieNo ratings yet
- The Chronobiology of Human Cytokines ProductionDocument15 pagesThe Chronobiology of Human Cytokines Productionيوميات صيدلانية pharmacist diariesNo ratings yet
- A New Portrait of Constitutive HeterochromatinDocument19 pagesA New Portrait of Constitutive HeterochromatinannyoryaNo ratings yet
- Triad Nihms 475933Document44 pagesTriad Nihms 475933poojaNo ratings yet
- Messengers From The MicrobiotaDocument3 pagesMessengers From The MicrobiotaDra. Ianna CarmoNo ratings yet
- Man Nello 2005Document15 pagesMan Nello 2005edelinNo ratings yet
- Ferroptosis: A Unique Form of Iron-Dependent Regulated Cell Death and Its Role in Different DiseasesDocument15 pagesFerroptosis: A Unique Form of Iron-Dependent Regulated Cell Death and Its Role in Different DiseasesSnehasis JanaNo ratings yet
- Virulence properties of the Legionella pneumophilaDocument12 pagesVirulence properties of the Legionella pneumophilaGuillermo RodríguezNo ratings yet
- The Role of Hepcidin in Iron MetabolismDocument10 pagesThe Role of Hepcidin in Iron MetabolismTatiana JalbaNo ratings yet
- Ferroptosis of Macrophages Facilitates Bone Loss in Apical Periodontitis Via NRF2 - FSP1 - ROS PathwayDocument14 pagesFerroptosis of Macrophages Facilitates Bone Loss in Apical Periodontitis Via NRF2 - FSP1 - ROS Pathwayratna saraswatiNo ratings yet
- Specs 5FE GBDocument5 pagesSpecs 5FE GBkemitaNo ratings yet
- Topnotch Surgery Supplement Handout UPDATED April 2016 PDFDocument83 pagesTopnotch Surgery Supplement Handout UPDATED April 2016 PDFDre ValdezNo ratings yet
- Susu HipoalergenikDocument14 pagesSusu HipoalergenikStella NoviaNo ratings yet
- Tumor Angiogenesis: Causes, Consequences, Challenges and OpportunitiesDocument26 pagesTumor Angiogenesis: Causes, Consequences, Challenges and OpportunitiesStella NoviaNo ratings yet
- Abnormal Vitamin Status in Childhood LeukemiaDocument8 pagesAbnormal Vitamin Status in Childhood LeukemiaStella NoviaNo ratings yet
- Effects of Vitamin B6 Metabolism On Oncogenesis, Tumor Progression and Therapeutic ResponsesDocument10 pagesEffects of Vitamin B6 Metabolism On Oncogenesis, Tumor Progression and Therapeutic ResponsesStella NoviaNo ratings yet
- Role of Central Vagal 5-HT Receptors in Gastrointestinal Physiology and PathophysiologyDocument10 pagesRole of Central Vagal 5-HT Receptors in Gastrointestinal Physiology and PathophysiologyStella NoviaNo ratings yet
- Free PPT Templates: Insert The Title of Your Presentation HereDocument3 pagesFree PPT Templates: Insert The Title of Your Presentation HereStella NoviaNo ratings yet
- Cancer Statistics, 2008Document26 pagesCancer Statistics, 2008Stella NoviaNo ratings yet
- Research ArticleDocument9 pagesResearch ArticleStella NoviaNo ratings yet
- Agenda Postgradmededu 120920 PDFDocument2 pagesAgenda Postgradmededu 120920 PDFStella NoviaNo ratings yet
- New England Journal Medicine: The ofDocument11 pagesNew England Journal Medicine: The ofStella NoviaNo ratings yet
- Nejmoa1911998 PDFDocument11 pagesNejmoa1911998 PDFStella NoviaNo ratings yet
- Free PPT Templates: PandemicDocument48 pagesFree PPT Templates: PandemicStella NoviaNo ratings yet
- Were The Babies Switched - Answer Sheet 2023 - GalvinDocument5 pagesWere The Babies Switched - Answer Sheet 2023 - Galvin25210580No ratings yet
- Reviewer (Abpsych)Document7 pagesReviewer (Abpsych)Joana Vivien Caraan100% (2)
- 2010-11 partA-F eDocument41 pages2010-11 partA-F eAltunc Suleyman GezerlerNo ratings yet
- Full Download Essentials of Physical Anthropology 10th Edition Jurmain Solutions ManualDocument35 pagesFull Download Essentials of Physical Anthropology 10th Edition Jurmain Solutions Manualudalbornhockeyspl2100% (31)
- Rheumatoid Arthritis: Pathophysiology PathophysiologyDocument34 pagesRheumatoid Arthritis: Pathophysiology PathophysiologyOmair RiazNo ratings yet
- Hormone 1Document14 pagesHormone 1IMDCBiochemNo ratings yet
- Spindle Cell Sarcomas: Cyril FisherDocument24 pagesSpindle Cell Sarcomas: Cyril FisherThierry GrimaNo ratings yet
- 1) Class 12th Biology Complete Unacademy BookDocument538 pages1) Class 12th Biology Complete Unacademy Bookkanikadudeja.07No ratings yet
- Biology Unit 3 Revision Year 8Document3 pagesBiology Unit 3 Revision Year 8Alan klnNo ratings yet
- PDF Diagnosis and Management of Mitochondrial Disorders Michelangelo Mancuso Ebook Full ChapterDocument53 pagesPDF Diagnosis and Management of Mitochondrial Disorders Michelangelo Mancuso Ebook Full Chapteredward.goodwin495100% (4)
- Detection of Malarial Parasite in Blood Using Image ProcessingDocument3 pagesDetection of Malarial Parasite in Blood Using Image ProcessingelectricalengineerNo ratings yet
- TA Brown Chapter 2 NotesDocument7 pagesTA Brown Chapter 2 Notesom prakashNo ratings yet
- File 1434571027Document1 pageFile 1434571027Guneyden GuneydenNo ratings yet
- Divine Intervention Episode 17 Diseases of The Pediatric Population Part 1Document14 pagesDivine Intervention Episode 17 Diseases of The Pediatric Population Part 1Swisskelly1No ratings yet
- Research Triangle Regions Life Sciences Directory 2012Document50 pagesResearch Triangle Regions Life Sciences Directory 2012hitexNo ratings yet
- The Immune System: HistoryDocument4 pagesThe Immune System: HistoryleesclassroomNo ratings yet
- A Project Proposal On Case Study and Management of A Type 1 Diabetes Mellitus ClientDocument6 pagesA Project Proposal On Case Study and Management of A Type 1 Diabetes Mellitus ClientMichael Kivumbi100% (3)
- An Update On Therapeutic Repurposing Strategies For COVID-19Document7 pagesAn Update On Therapeutic Repurposing Strategies For COVID-19Prakash KatakamNo ratings yet
- Listening Sample Test 1 Audio ScriptDocument11 pagesListening Sample Test 1 Audio ScriptCCCAXANo ratings yet
- Principles of Drug Addiction Treatment A Research Based Guide Third EditionDocument67 pagesPrinciples of Drug Addiction Treatment A Research Based Guide Third EditionКонстантин Крахмалев100% (1)
- Ankylosing Spondylitis (Marie-Strümpell Disease/ Bechterew's Disease)Document39 pagesAnkylosing Spondylitis (Marie-Strümpell Disease/ Bechterew's Disease)MoineeNo ratings yet
- Mod 2 Comp 13 - Demonstrate Knowledge of Electrical Safety and Safe Working Practices For Electrical WorkersDocument9 pagesMod 2 Comp 13 - Demonstrate Knowledge of Electrical Safety and Safe Working Practices For Electrical WorkersSmr OnlyNo ratings yet
- Biology - Form 2 Scheme of Work - Term 1: Week Topic Syllabus Objectives Number OF Sessions Activities/LabsDocument2 pagesBiology - Form 2 Scheme of Work - Term 1: Week Topic Syllabus Objectives Number OF Sessions Activities/LabsramjdNo ratings yet
- Juvenile Nasopharyngeal AngiofibromaDocument28 pagesJuvenile Nasopharyngeal AngiofibromamaNo ratings yet
- Health Benefits of Coconut Milk23Document6 pagesHealth Benefits of Coconut Milk23Gerson Paul BangoyNo ratings yet
- Premalignant LesionsDocument76 pagesPremalignant LesionsPrima D Andri100% (1)
- Small Animal OncologyDocument103 pagesSmall Animal Oncologyvet53No ratings yet